





Abstract
Fibroblasts play an important role in the repair and remodelling processes following injury. Prostaglandin D2 (PGD2) is a potent mediator in inflammatory processes.
In this study, the effect of the PGD2 on human foetal lung fibroblasts (HFL‐1) chemotaxis induced by human plasma fibronectin (HFn) was investigated using the blindwell chamber technique.
PGD2 inhibited HFL‐1 chemotaxis to HFn (20 µg·mL−1) by 20.8±3.8% (p<0.05). Checkerboard analysis of HFn-directed migration confirmed that PGD2 inhibited both chemotaxis and chemokinesis. The effect of PGD2 was concentration-dependent and the inhibitory effect diminished with time. The PGD2 receptor (DP) agonist BW245C (500 nM) had a similar effect, inhibiting chemotaxis to 39.4±6.3%. The inhibitory effects of both PGD2 and BW245C on HFL‐1 chemotaxis were blocked by the DP receptor antagonist AH6809 (2 µM). The inhibitory effect of PGD2 on fibroblast chemotaxis was also blocked by the cyclic adenosine monophosphate (cAMP)‐dependent protein kinase (PKA) inhibitor, KT5720, suggesting a DP receptor-initiated, cAMP‐dependent effect mediated by PKA.
Prostaglandin D2 appears to inhibit fibroblast chemotaxis, perhaps by modulating the rate of fibroblast migration. Such an effect may contribute to regulation of the wound healing response following injury in asthma patients.
This study was funded by the Larson Endowment, University of Nebraska Medical Centre and NIH grant #HL64088‐03.
Under normal circumstances, tissue structure and function is restored after acute inflammation caused by injury. The migration of fibroblasts from neighbouring connective tissues to sites of inflammation plays an important role in tissue repair. If inadequate fibroblast recruitment is present, repair may be defective. In contrast, excessive accumulation of fibroblasts can result in fibrosis with alteration in tissue architecture and loss of function 1–4.
Bronchial asthma is characterized by bronchospasm, oedema and hypersecretion. Mast cells are believed to play an important role in acute asthma by secreting a number of mediators including histamine, prostaglandin D2 (PGD2) and cysteinyl leukotrienes 5, 6. Moreover, mast cells may be involved in airway remodelling since they have been found to play an important part in pulmonary fibrosis and also secrete mediators such as transforming growth factor‐β, which can modulate repair and remodelling 7, 8. Asthma is also characterized by remodelling of the airway walls, a prominent feature of which is the accumulation of mesenchymal cells and deposition of extracellular matrix. The mediators responsible for this process remain to be defined.
The current study was designed to evaluate the hypothesis that PGD2, in addition to contributing to acute symptoms in asthma, might also be able to contribute to remodelling by modulating fibroblast recruitment. PGD2 is the most potent bronchoconstricting prostanoid known and is 30‐times stronger than histamine as a bronchoconstricting agent in asthmatic subjects with a longer duration of action 9. PGD2 is a product of the cyclo-oxygenase pathway of arachidonic acid metabolism, and is primarily derived from prostaglandin H2. In addition to mast cells, PGD2 is synthesized by alveolar macrophages and platelets, and produces a variety of physiological responses, including vascular permeability elevation, vascular relaxation 10, platelet diffusion 11, and sleep induction. The biological actions of PGD2 are mediated by DP class prostanoid receptors that belong to the superfamily of specific G‐protein-coupled receptors 12. The DP receptor couples with a G‐protein to trigger adenylyl cyclase, which, in turn, catalyzes the hydrolysis reaction of adenosine triphosphate (ATP) to the second messenger cyclic adenosine monophosphate (cAMP) 12. The current study was designed to evaluate the effect of PGD2 on fibroblast chemotaxis, using purified human plasma fibronectin (HFn) as a chemoattractant. In addition, the mechanism by which PGD2 exerts its effect on fibroblast chemotaxis was evaluated.
Materials and methods
Materials
The PGD2, DP receptor agonist BW245C, receptor antagonist AH6809, and thromboxane A2 (TP) receptor antagonist SQ29,548 were purchased from Cayman Chemical (Ann Arbor, MI, USA). KT5720 was purchased from Calbiochem (San Diego, CA, USA). PGD2 and KT5720 were dissolved in DMSO at 1×10−2 M. BW245C (1×10−2 M), AH6809 (1×10−3 M) and SQ29,548 (2×10−3 M) were dissolved in ethanol. Tissue culture supplements and media were purchased from GIBCO (Life Technologies, Grand Island, NY, USA). Foetal calf serum (FCS) was purchased from Biofluid (Rockville, MD, USA).
Human foetal lung fibroblasts
Human foetal lung fibroblasts (HFL‐1) were obtained from the American Type Culture Collection (Rockville, MD, USA). The cells were cultured in 100 mm tissue culture dishes (Falcon; Becton-Dickinson Labware, Lincoln Park, NJ, USA) in Dulbecco's modified Eagle medium (DMEM, Gibco, Grand Island, NY, USA), supplemented with 10% FCS, 50 U·mL−1 penicillin G sodium, 50 µg·mL−1 streptomycin sulphate (penicillin-streptomycin, GIBCO), and 1 µg·mL−1 amphotericin‐B (Parma-Tek, Huntington, NY, USA) in a humidified atmosphere at 37°C and 5% CO2. The fibroblasts were routinely passaged every 4–5 days and cells were used between passage 13 and 20 in all experiments. Confluent fibroblasts were removed from the dishes by treatment with 0.05% trypsin in 0.53 mM ethylenediamine tetraacetic acid and resuspended in DMEM without serum.
Human fibronectin
HFn was purified from human plasma by a modification of the affinity chromatography method described previously 13, 14. Briefly, after passage through a Sepharose 4B column to eliminate nonspecific absorption, affinity chromatography was performed on gelatin sepharose 4B followed by affinity chromatography on heparin agarose.
Fibroblast chemotaxis
HFL‐1 chemotaxis was assessed by the Boyden blindwell chamber technique 15 using 48‐well chambers (Nucleopore, Cabin John, MD, USA). The chemoattractant, i.e. HFn, was placed in the bottom chamber. In some experiments, PGD2 was also added to the lower chamber. An 8 µm pore filter (Nucleopore, Pleasanton, CA, USA) coated with 0.1% gelatin (Bio Rad, Hercules, CA, USA) was placed over the lower portions of the chamber. The top manifold was placed and HFL‐1 (1.0×106 mL in DMEM without serum) were loaded into the upper well of the chamber with the desired concentration of PGD2 or other additives. The chamber was then incubated at 37°C in a moist, 5% CO2 atmosphere. Except as designated, chambers were incubated for 6 h, after which the cells on the top of the filter were removed by scraping. The filter was then fixed, stained with Protocol (Biochemical Science, Swedesboro, NJ, USA), and mounted on a glass microscope slide. Migration was assessed by counting the number of cells in five high-power fields using a light microscope. Triplicate wells were prepared in each experiment for every condition. Replica experiments were performed with separate cultures of cells on separate occasions.
Statistical analysis
Samples with multiple comparisons were analysed for significance using analysis of variance (ANOVA). Where ANOVA indicated significant differences between groups, for the preplanned comparisons of interest, the Tukey correction was applied and p<0.05 was taken as significant. Summary data are expressed as mean±sem.
Results
As expected, HFn‐directed HFL‐1 chemotaxis occurred in a concentration-dependent manner (fig. 1⇓).

Chemotaxis of human foetal lung fibroblast using purified human plasma fibronectin (HFn) (□: control) and with the addition of prostaglandin D2 (1×10−6 M: ⋄). Data are expressed as mean±sem. *: p<0.05.
PGD2 (1×10−6 M), when added to the fibroblasts immediately before the cells were placed in the top wells of the chemotaxis chamber, inhibited HFn‐directed chemotaxis with various HFn concentrations (≥20 µg·mL−1) (fig. 1⇑). This result was consistently observed. Similar results were observed with platlet derived growth factor (PDGF)‐BB (10 ng·mL−1) as chemoattractant (data not shown). In three separate experiments, 1×10−6 M of PGD2 inhibited fibroblast chemotaxis to 20 µg·mL−1 HFn by 20.8±3.8% (p<0.05). The inhibitory effect of PGD2 was concentration-dependent (fig. 2⇓).

Concentration-dependent inhibition of human foetal lung fibroblast chemotaxis by prostaglandin D2 (PGD2). Data are expressed as mean±sem for three sample cultures of fibroblasts exposed to PGD2 on different days. *: p<0.05
The number of HFL‐1 that accumulated on the bottom of the chemotaxis chamber increased as a function of time, both for control HFL‐1 and for those treated with PGD2 (fig. 3⇓). Control HFL‐1 chemotaxis was near maximal by 12 h. In contrast, the number of migrated HFL‐1 exposed to PGD2 was still increasing at 24 h, the last time point evaluated. The differences between PGD2 and control, which were readily observed at 6 h, were not statistically significant at later time points. Thus, it would appear that PGD2 has a greater effect on the rate of fibroblast migration than on the number of migrating cells.

Time course of the inhibition of human foetal lung fibroblast chemotaxis by prostaglandin D2 (PGD2). Data are presented as mean±sem. □: control; ⋄: 1×10−7 M PGD2; ○: 1×10−6 PGD2. *: p<0.05.
To determine if PGD2 inhibited chemotaxis, chemokinesis or both, varying concentrations of HFn were placed both above and below the filter. This method allowed migration to be measured in the presence of increasing concentrations, but in the absence of a gradient (chemokinesis), as shown in table 1⇓, and in the presence of a gradient (chemotaxis), as shown by the vertical columns. The number of cells migrating increased as the concentration of HFn increased in the absence of a gradient (diagonal) indicating that chemokinesis was present. Similarly, the number of migrated cells increased when a gradient was present, indicating chemotaxis was also present. PGD2 inhibited both chemotaxis and chemokinesis in a concentration-dependent manner.
Demonstrates whether prostaglandin D2 (PGD2) inhibits chemotaxis, chemokinesis or both at ranging concentration of human fibronectin (HFn)
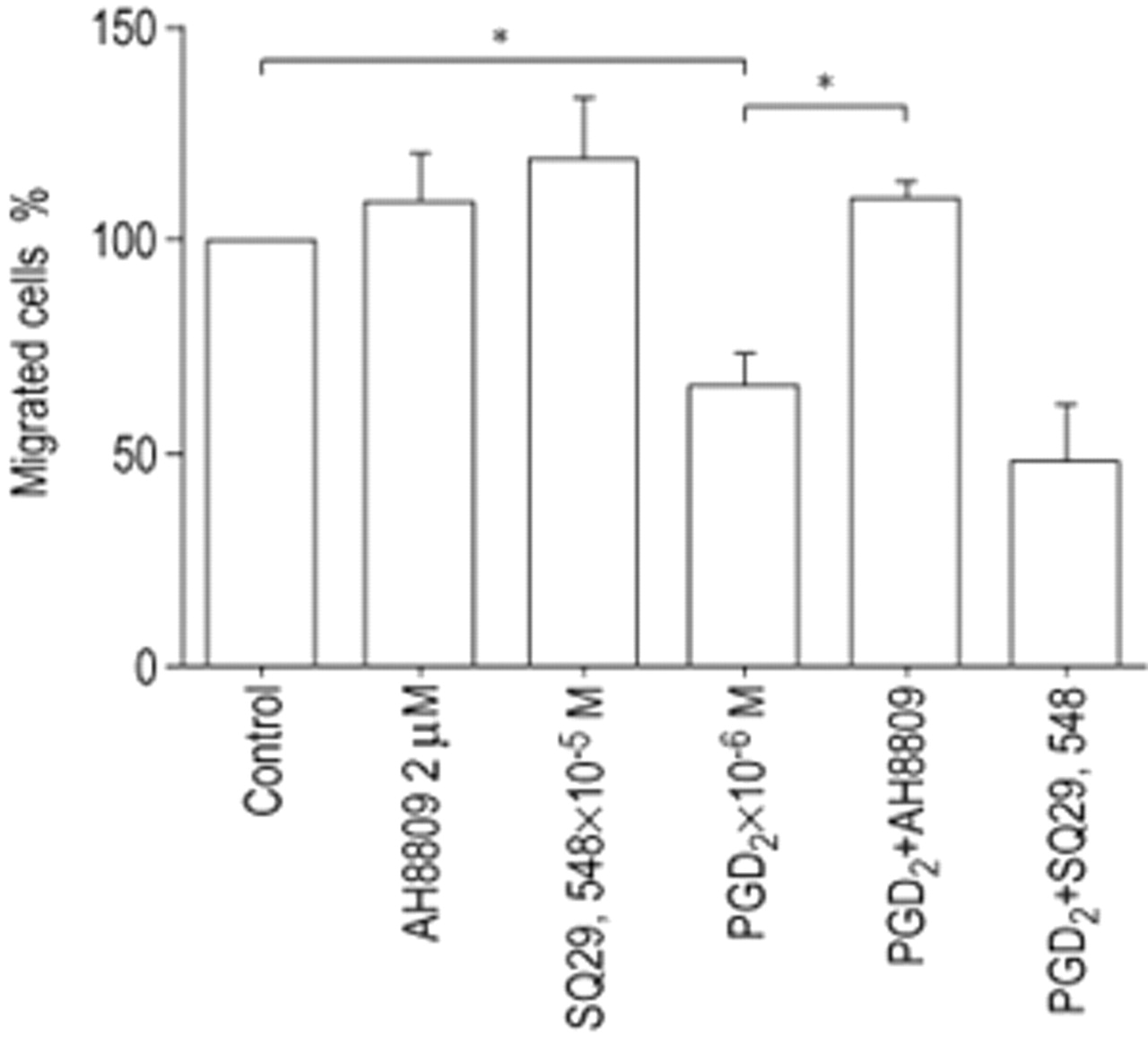
To determine if the inhibitory effect of PGD2 is mediated through the DP receptor, HFL‐1 cells were treated with the inhibitor AH6809 (2 µM) for 1 h before harvesting for the chemotaxis assay. AH6809 blocked the PGD2‐mediated inhibition of fibroblast chemotaxis to HFn (fig. 4⇓). In contrast, the TP receptor antagonist SQ29,548 had no effect on PGD2‐mediated inhibition of fibroblast chemotaxis.

Effect of prostaglandin D2 (PGD2) or thromboxane A2 (TXA2) receptor antagonists on inhibition of human foetal lung fibroblast chemotaxis by PGD2. AH6809: PGD2 receptor antagonist; SQ29,548: TXA2 receptor anatogonist. Data are presented as mean±sem for three separate experiments. *: p<0.05
To further define the role of the DP receptor in mediating PGD2 inhibition of fibroblast chemotaxis, the DP receptor agonist BW245C was added to fibroblasts for 1h and was observed to cause a concentration-dependent inhibition of fibroblast chemotaxis (fig. 5⇓).

Inhibition of human foetal lung fibroblast chemotaxis by BW245C with (○) or without (□) AH6809. Data are presented as the mean±sem of three separate experiments. BW245C: prostaglandin (PGD2) receptor agonist; AH6809: PGD2 receptor antagonist. *: p<0.05.
The DP receptor is known to couple to adenylyl cyclase and increase cAMP 16, and other agonists are known to inhibit fibroblast chemotaxis through cAMP 17. To confirm a cAMP‐mediated inhibition of fibroblast chemotaxis by PGD2, the protein kinase A (PKA) inhibitor KT5720 was used. Preincubation with KT5720 (1×10−7 M) in monolayer culture for 1 h caused a rightward shift in the PGD2 inhibition curve, consistent with its competitive antagonism with cAMP for PKA activation (fig. 6⇓).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of inhibition of protein kinase A (PKA) on prostaglandin D2 (PGD2) inhibition of human foetal lung fibroblast chemotaxis to human plasma fibronection. Data are presented as the mean±sem of three separate experiments. □: control; ○: 1×10−7 M KT5720 (PKA inhibitor). *: p<0.05.
Discussion
The current study demonstrates that PGD2 is capable of inhibiting fibroblast chemotaxis to purified HFn in a concentration-dependent manner. Cell migration continued over time suggesting that the effect of PGD2 was to decrease the rate of migration rather than the number of cells migrating. Both chemotaxis and chemokinesis were affected. The PGD2 receptor agonist BW245C demonstrated the same effect as PGD2 on fibroblast chemotaxis, and the inhibitory effect on fibroblast chemotaxis was blocked by the DP receptor antagonist AH6809 but not by the TP receptor antagonist SQ29548, indicating that the inhibitory effect on fibroblast chemotaxis was mediated through the DP receptor. Consistent with this and the known signalling of the DP receptor through cAMP, the effect of PGD2 was antagonized by the competitive inhibitor of PKA, KT5720.
Alteration in tissue structure is a characteristic feature of asthma and other allergic diseases. In asthma, mesenchymal cells accumulate in the subepithelial tissue and are thought to be responsible for producing extracellular matrix macromolecules 18, 19, thereby making them major contributors to alterations in tissue structure. The mechanisms by which these mesenchymal cells accumulate are likely to be multifactorial and involve both the chemotactic recruitment of cells to sites of inflammation, as well as expansion of cell numbers in response to specific growth factors 20. Overall tissue balance probably also depends on removal of cells through a variety of mechanisms including factors which drive apoptosis. The ability of PGD2 to inhibit chemotaxis is not specific for HFn as evidenced by its ability to inhibit PDGF‐BB chemotaxis. Chemotaxis depends on at least two cellular functions 21. First, cellular receptors must recognize a concentration gradient in the chemoattractant. Second, the cell must be able to migrate in response to the chemoattractant signal. Inhibition of chemotaxis can result from either inhibition in the recognition of the chemotactic gradient or from inhibition of the ability of the cell to migrate. The “checkerboard” analysis in the current study suggests that PDG2 inhibits the ability of the cells to migrate.
The current study supports the concept that some mediators present in an inflammatory milieu could also serve to restrict mesenchymal cell accumulation by inhibiting fibroblast recruitment. In this context, PGD2 is a mediator characteristically present in inflamed tissues in asthma. Mast cells are believed to be a prominent source of PGD2 5, 22, and increased levels of PGD2 have been reported in lavage fluid from asthmatics 23. Other prostaglandins, for example prostaglandin E2 (PGE2), also have been shown to modulate mesenchymal cell chemotaxis 24. Interestingly, PGE2 can inhibit fibroblast chemotaxis but accelerates epithelial cell wound closure, suggesting that prostanoids may exert a variety of effects on cells participating in wound repair 25.
Prostaglandins exert their effects on G‐protein coupled receptors. At least 10 receptors are involved in prostaglandin signalling, and many prostaglandins can interact with multiple receptors. In this regard, PGD2 has been reported to interact with both the DP receptor and the TP receptor 26. The DP receptor is believed to activate adenylyl cyclase and lead to an increase in cAMP 17. The TP receptor, in contrast, is believed to activate phospholipase C leading to generation of inositol 1, 4, 5‐triphosphate with activation of protein kinase C and leading to subsequent changes in intracellular calcium 27.
Interestingly, the bronchoconstrictory effect exerted by PGD2 is believed to be mediated through the TP receptor 26. Hirai et al. 28, recently demonstrated that PGD2 could induce chemotaxis of T‐helper 2 cells, eosinophils and basophils by way of the seven-transmembrane receptor CRTH2. In the current study, in contrast, the ability of PGD2 to inhibit fibroblast chemotaxis appears to be mediated through the DP receptor and not through the TP receptor. In this context, the effect of PGD2 is similar to other agents that also inhibit fibroblast chemotaxis by increasing intracellular cAMP levels 24.
Many mediators can regulate fibroblast recruitment and accumulation. Maintenance of tissue structure probably depends on a dynamic balance between factors that stimulate and inhibit these processes. By contributing to this balance, prostaglandin D2 can play a role not only in the acute events initiated by allergens, for example vascular leak and bronchoconstriction, but also in tissue remodelling.
Acknowledgments
The authors are pleased to acknowledge the support of L. Richards and M. Tourek for assistance with preparation of this manuscript.
- Received July 9, 2001.
- Accepted November 7, 2001.
- © ERS Journals Ltd
References









