





Abstract
In acute lung injury, a decrease in surface-active large aggregates and an increase in the less surface-active small surfactant aggregates are observed. The objective of the current study was to determine if the increase in small aggregates interfered with the function of large aggregates, thereby independently contributing to lung dysfunction.
Isolated large aggregates, small aggregates, and large aggregate+small aggregate combinations were analysed for in vitro surface activity utilizing a pulsating bubble surfactometer. Subsequently, large aggregates, small aggregates, and large aggregate+small aggregate combinations were administered to surfactant-deficient, adult Sprague-Dawley rats. Physiological parameters were measured during 1 h of ventilation. After sacrifice, the whole lung lavage was analysed for protein concentration, and surface activity of the recovered large aggregates.
The minimum surface tension of the large aggregate+small aggregate preparations (10 mN·m−1) was significantly higher than large aggregates alone (1 mN·m−1), but lower than small aggregates alone (21 mN·m−1) after 100 pulsations. In vivo, rats receiving large aggregates+small aggregates showed immediate increases in oxygenation, similar to animals given large aggregates, whereas animals given small aggregates and control animals maintained low oxygenation values.
In conclusion, small aggregates interfered with large aggregates function in vitro, but this was not observed in vivo in this experimental model.
Pulmonary surfactant can be separated into two subfractions via centrifugation; dense structures representing the surface-active subfraction of alveolar surfactant termed “large aggregates” (LA), and less surface-active, lighter vesicular forms called “small aggregates” (SA) 1. LA are responsible for maintaining alveolar stability through the reduction of surface tension arising at the air-liquid interface of the alveoli. During respiration, LA are converted into SA, which are either recycled back into the type II cell or cleared from the airspace 2.
Within the airspace of normal, healthy lungs, a consistent proportion of LA and SA exist 1. However, in acute lung injury, and particularly its more severe form, the acute respiratory distress syndrome (ARDS), several alterations of the surfactant system are evident, including changes in the relative amounts of these aggregate forms 1, 3, 4. Studies utilizing animal models of ARDS have reported increased SA forms and decreased LA forms with, in some cases, no significant change in total alveolar surfactant pool sizes 3, 5, 6. A relative increase in SA has also been demonstrated in bronchoalveolar lavage samples obtained from patients with ARDS 4, 7. Although these changes in aggregate forms have been shown to contribute to lung dysfunction, it is unknown to what extent the increase in SA contributes to lung injury, independent of the decrease in functionally active LA. The objective of the current study was to determine if the presence of SA interferes with the function of LA.
Methods
Animals
Thirty-six adult male Sprague-Dawley rats (Charles River, Constant, PQ, Canada), averaging 374±4.0 g, were used for in vivo aggregate analysis. Experimental procedures were approved by The University of Western Ontario Animal Care Committee in accordance with Canadian Council of Animal Care guidelines.
Study design
To determine whether SA interfered with LA function, both in vitro and in vivo experiments were performed. The in vitro experiments involved analysis of the surface tension-reducing ability of LA and SA, as well as mixtures of these two subfractions, using a pulsating bubble surfactometer. For in vivo analyses, LA, SA, and mixtures of LA and SA were instilled into the lungs of surfactant-deficient adult rats. Animals were ventilated for 1 h, monitoring physiological parameters. Subsequently, lungs were lavaged for quantification of total protein and assessment of the surface activity of recovered LA.
Surfactant aggregate preparations
Surfactant aggregates were isolated from whole lung lavage obtained from healthy adult sheep using 0.15 M NaCl. The lavage was centrifuged at 150×g to remove cellular debris, the supernatant from this centrifugation was then spun at 40,000×g for 15 min at 4°C to obtain a pellet corresponding to the LA fraction 1. Pelleted material was resuspended in sterile 0.15 M NaCl, pooled, and further purified by sucrose gradient centrifugation 8. The resulting pellet was resuspended in sterile 0.15 M NaCl, centrifuged again for 15 min at 40,000×g to remove remaining sucrose, and resuspended and frozen at −20°C until further use.
The supernatant from the initial 40,000×g spin contained the SA fraction 1. This supernatant was concentrated using an ultrafiltration system equipped with 300,000 kDa retention filters (Millipore, Bedford, MA, USA), pooled, and subsequently dried via refrigerated speed vac (Savant Instruments Inc., Farmingdale, NY, USA). SA were resuspended in sterile 0.15 M NaCl, and organic solvent-extracted using chloroform-methanol 9. The dried sample was resuspended in sterile 0.15 M NaCl and frozen at −20°C until further use. In experiments performed to assess SA uptake, these preparations were radiolabelled with 3H-dipalmitoylphosphatidylcholine (DPPC) as previously described 5.
Analyses of surfactant aggregate preparations
A phosphorus assay to determine total phospholipid-phosphorus was performed on the purified LA and SA by a modification of the Duck-Chong method 10. Phospholipid composition was analysed by separating the various phospholipids using thin-layer chromatography 6. Individual phospholipid species were subsequently quantified using the method of Duck-Chong 10. Morphological analyses via electron microscopy were also performed as previously described 11. The in vitro surface activity of the LA and SA samples, both alone and in combination, was assessed using a pulsating bubble surfactometer (Electronetics, Buffalo, NY, USA) 12. Aliquots of the LA and SA were resuspended in 0.15 M NaCl/1.5 mM CaCl2 to concentrations of 1 mg and 2 mg phospholipid·mL−1. Two different mixtures of LA and SA (1 mg phospholipid·mL−1 LA with 1 mg·mL−1 SA, and 1 mg·mL−1 LA with 2 mg·mL−1 SA) were also prepared. With this technique, an air bubble is formed in a surfactant suspension and pulsated at a rate of 20 pulsations·min−1 between a maximum and minimum radius of 0.55 and 0.4 mm respectively, with temperature maintained at 37°C. Values are expressed as surface tension in mN·m−1 following 10 s of adsorption, and 100 pulsations at minimum bubble radius.
Saline lavage rat model
Rats were anaesthetized with an intraperitoneal injection of ketamine hydrochloride (75 mg·kg−1 body weight) and xylazine (5 mg·kg−1). The jugular vein was cannulated using polyethylene tubing (PE-50) to allow for saline and anaesthetic infusion. Similarly, the carotid artery was cannulated for blood gas measurements and arterial blood pressure monitoring. A tracheotomy was performed by inserting a 14 gauge angiocatheter as an endotracheal tube. Animals were anaesthetized with sodium pentabarbitol (20–40 mg·kg−1), and paralysed with pancuronium bromide (2 mg·kg−1) to inhibit spontaneous breathing. Animals were subsequently connected to a volume-cycled rodent ventilator (Harvard Instruments, Saint Laurent, PQ, Canada) with the following parameters: a fraction of inspired oxygen (FI,O2) of 1.0, respiratory rate of 73 breaths·min−1, tidal volume of 8 mL·kg−1, and a positive end-expiratory pressure of 5 cmH2O. Peak inspiratory pressure (PIP) and arterial blood gases were measured (ABL-500, Radiometer, Copenhagen, Denmark) after a 10 min stabilization period, representing baseline values.
Surfactant-deficiency was then induced via a repetitive whole lung saline lavage procedure by slowly infusing 10 mL of warmed, sterile 0.15 M NaCl into the lungs and gently withdrawing. This procedure was repeated three times, at 5 min intervals. Arterial blood gases were analysed following the fourth lavage, and subsequent lavages were performed at 10 min intervals thereafter, until the oxygen tension in arterial blood (Pa,O2)/(FI,O2) values ranged between 60–100 mmHg. Preliminary studies showed that these criteria ensured animals were surfactant-deficient and stable for the remainder of the experiment.
After the final lavage, animals were randomized into one of the following six experimental groups (n=6·group−1): 1) 15 mg phospholipid·kg−1 body weight LA (15 LA); 2) 15 mg·kg−1 SA (15 SA); 3) 30 mg·kg−1 SA (30 SA); 4) 15 mg·kg−1 LA+15 mg·kg−1 SA (15 LA/15 SA); 5) 15 mg·kg−1 LA+30 mg·kg−1 SA (15 LA/30 SA), or 6) air bolus alone (control). A seventh group (30 mg·kg−1 LA) was also included in the randomization, however, since the physiological and biophysical results obtained with this group were similar to the 15 mg·kg−1 LA group, this data was not included in this report.
Animals were disconnected from the ventilator and the surfactant preparation was instilled from a syringe into the endotracheal tube, followed by an air bolus of 10 mL·kg−1 to optimize peripheral distribution. The syringe was disconnected after maintaining lung inflation via the air bolus for 10 s, and animals were reconnected to the ventilator. The concentrations of aggregates in the various groups were adjusted so that all animals received the same instilled volume of 4 mL·kg−1. This ensured similar surfactant distribution patterns within the lungs after administration of the various doses 13. Blood gases and PIP values were monitored at 5, 15, 30, 45, and 60 min post-surfactant instillation.
Following the 60 min ventilatory period, animals were sacrificed by an intravascular overdose of sodium pentabarbitol (>100 mg·kg−1) and exsanguinated via transection of the descending aorta. The lungs were visualized and a whole lung lavage procedure was performed and processed to obtain the LA and SA as previously described 14. Surfactant phospholipid-phosphorus content was assessed utilizing the method of Duck-Chong 10, total protein was measured in lavage samples using a Lowry protein assay 15, and surface activity of the recovered LA fractions (1 mg phospholipid·mL−1) was assessed using the pulsating bubble surfactometer.
In experiments in which radioactive SA were administered (in the 15 and 30 mg·kg−1 SA groups), radioactivity recovered in the lavage fluid and homogenized lung tissue was analysed via liquid scintillation counting, after aliquots were organic-solvent extracted and dried. This data was expressed as radioactivity recovered in the lavage as a per cent of total radiolabel recovery (tissue + lavage).
Statistical analyses
All data are expressed as means±standard error of the mean (sem). Comparisons within and among the experimental groups were made using an analysis of variance (ANOVA) followed by a Bonferroni post-hoc test. A p-value <0.05 was considered significant.
Results
Analyses of surfactant aggregate preparations in vitro
Table 1⇓ shows the phospholipid composition of the LA and SA preparations utilized in this study. Compositions were similar between the subfractions, although SA tended to have a slightly higher percentage of lysophosphatidylcholine than LA. Morphological analyses via electron microscopy showed that LA had organized lipid protein structures, such as lamellar bodies and tubular myelin as reported previously 11. These organized structures were not present in the SA preparations, which revealed lipid vesicular forms, with no distinct structural integrity. Figure 1⇓ shows the in vitro surface activity of the purified aggregate forms. After the 10 s adsorption period, surface tension values were approximately 29 mN·m−1 in the SA samples (1 and 2 mg·mL−1) and remained above 20 mN·m−1 over 100 pulsations, with no significant differences between the two concentrations tested. LA (1 and 2 mg·mL−1) had surface tension values of approximately 24 mN·m−1 after 10 s adsorption, and values decreased to near 0 mN·m−1 over 100 pulsations, which was significantly lower than both SA samples (p<0.001). When SA were combined with LA, surface tension values at adsorption were similar to those obtained for the LA fractions, however over 100 pulsations, values decreased to approximately 10 mN·m−1. This was significantly higher than LA (p<0.001), and significantly lower than SA alone (p<0.001). There were no significant differences between the two combinations of aggregate samples analysed.

Mean surface tension values of the purified aggregate preparations, in various concentrations (1 and 2 mg phospholipid·mL−1) as analysed via the pulsating bubble surfactometer (n= 4 samples·group−1). Small aggregates (SA) at both concentrations (▾: 1 mg·mL−1 SA; ▿: 2 mg·mL−1 SA) had high surface tension values over 100 pulsations. Large aggregates (LA) showed a rapid decrease in surface tension following adsorption (•: 1 mg·mL−1 LA; ○: 2 mg·mL−1 LA), while the LA+SA combinations (▪: 1 mg·mL−1 LA+1 mg·mL−1 SA; □: 1 mg·mL−1 LA+2 mg·mL−1 SA) showed only a slight decrease after adsorption over 100 pulsations. Statistical analysis only displayed after 20 pulsations. Error bars have been obscured where the range of SE falls within the symbol. #: p<0.05 SA (both) versus LA, LA+SA groups; ¶: p<0.05 LA+SA (both) versus SA, LA groups, +: p<0.05 LA (both) versus SA, LA+SA groups.
Phospholipid composition of the purified large aggregates (LA) and small aggregates (SA) (15 µg phosphorus) based on thin-layer chromatography expressed as a percentage of total phosphorus
Surfactant aggregate analyses in vivo
Figure 2⇓ shows the oxygenation values of animals receiving either surfactant aggregates or an air bolus (control), and ventilated for 1 h. In the control animals, Pa,O2/FI,O2 values remained low during the ventilation period (fig. 2a⇓). Animals given the 15 and 30 mg·kg−1 doses of SA had low oxygenation values, which were not significantly different from the values in the control animals over time. In contrast, animals administered the LA preparation (15 mg·kg−1) showed an immediate and significant increase in oxygenation (p<0.001 versus Time 0). These values were sustained throughout the ventilatory period. Pa,O2/FI,O2 values in the LA group were significantly higher than values measured in animals receiving either dose of SA, as well as the control group at all time points (p<0.01). Figure 2b⇓ shows the oxygenation values for animals receiving the combinations of LA and SA. Significant improvements in oxygenation were observed for both groups of animals immediately after administration. These values remained significantly higher than their respective pre-instillation values over the subsequent period of ventilation (p<0.001), and not significantly different from values recorded for animals receiving LA alone, but higher than both SA groups and the control group (p<0.005).
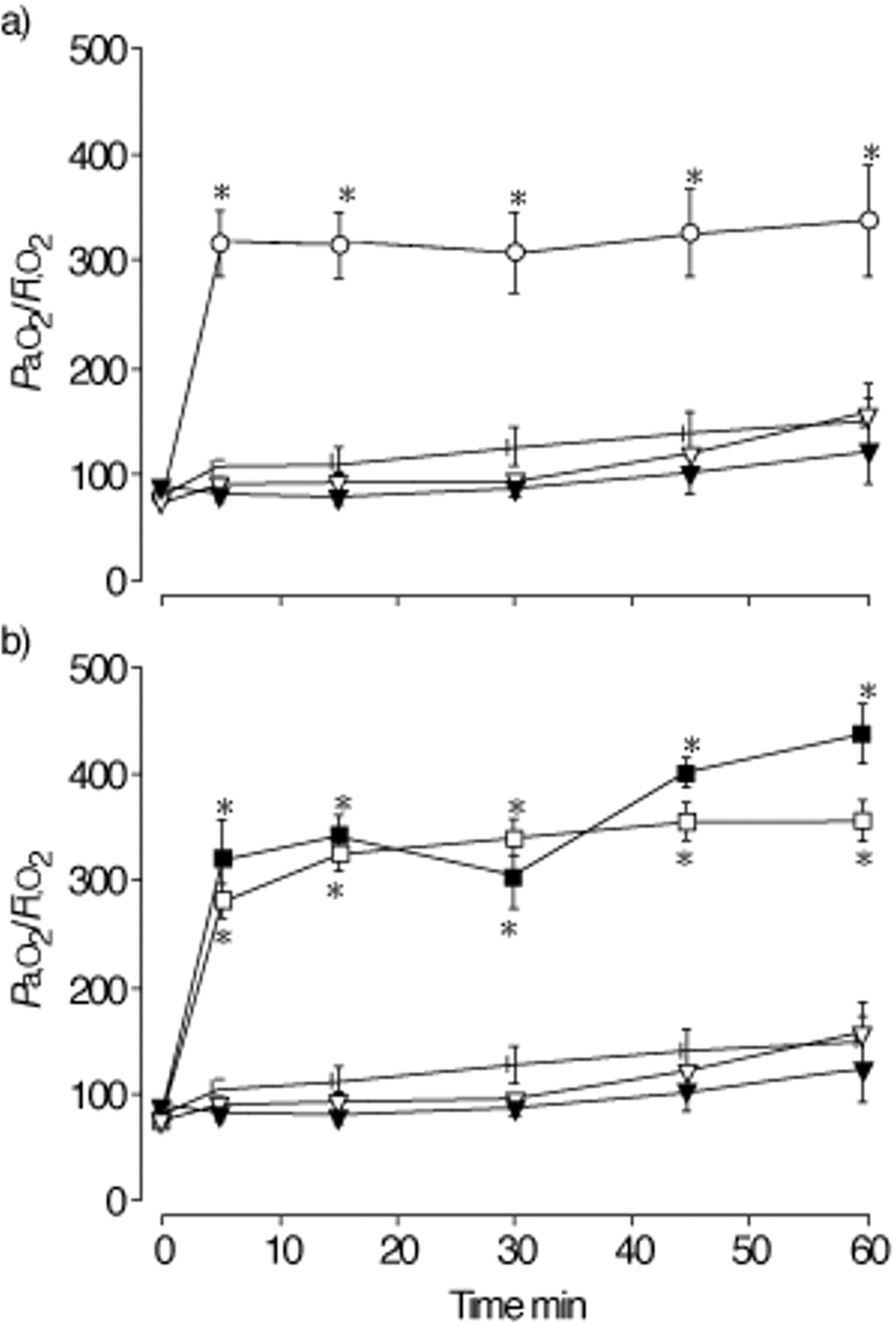
Mean oxygen tension in arterial blood (Pa,O2)/inspiratory oxygen fraction (FI,O2) values following administration of aggregates (or air bolus) at Time 0 (n=6 animals·group−1). a) The control (•) and small aggregate group (SA) (▾: 15 SA, ▿: 30 SA) animals had low oxygenation values with no improvement over the ventilatory period. The large aggregate group (LA) animals (○: 15 LA) showed a significant increase in oxygenation remaining consistent after aggregate instillation. *: p<0.05 versus control and SA groups. b) Animals receiving LA+SA combination at either concentration (▪: 15 LA/15 SA; □: 15 LA/30 SA) showed a significant increase in oxygenation, similar to the LA group animals. *: p<0.05 versus control, SA groups.
PIP values are shown in figure 3⇓. PIP values for the control group were greater than 19 cmH2O and remained consistent over the 60 min period of ventilation (fig. 3a⇓). Animals receiving both doses of SA had PIP values of approximately 20 cmH2O over the 60 min, which were similar to their respective pre-instillation values (Time 0), and not significantly different from the control group. In animals given the LA, PIP values decreased significantly by the 5 min time point (p<0.05) and remained consistent over the subsequent 60 min. These values were significantly lower than PIP values in both SA groups (p<0.05). After administering preparations containing the combination of aggregate forms (Figure 3b⇓), PIP values decreased significantly for both groups compared to their respective pre-instillation values (p<0.05), and remained consistent at approximately 16 cmH2O throughout the ventilatory period. These values did not differ significantly from PIP values recorded in the LA group, but were significantly lower than PIP values observed in both SA groups (p<0.05). There were no significant differences in PIP values between the groups receiving 15 LA/15 SA and 15 LA/30 SA.

{kind=link}
{kind=link}
{kind=link}
Mean peak inspiratory pressure (PIP) values following aggregate instillation (or air bolus) at Time 0 (n=6 animals·group−1). a) Control (•) animals had high PIP values, consistent over 1 h of ventilation. Small aggregate (SA) group animals (▾: 15 SA; ▿: 30 SA) were similar to control animals, while animals receiving large aggregates (LA) (○: 15 LA) showed a significant decrease in PIP following administration. *: p<0.05 versus control, SA groups; #: p<0.05 versus 15 SA group; ¶: p<0.05 versus SA groups. b) Animals receiving LA+SA combinations (▪: 15 LA/15 SA; □: 15 LA/30 SA) showed a significant decrease in PIP, similar to the LA group. *: p<0.05 versus control, SA groups; ¶: p<0.05 versus SA groups.
Analysis of the lung lavage fluid obtained after sacrifice following 60 min of ventilation showed similar quantities of total protein recovered from all groups (table 2⇓). Analysis of the surface activity of the recovered LA, indicated that the LA fraction isolated from animals receiving the SA (15 SA, 30 SA), had surface tension values of approximately 45 mN·m−1 after adsorption, but decreased significantly after 100 pulsations to values of 7.4±1.4 and 7.7±1.0 mN·m−1 respectively (table 2⇓). Samples obtained from animals receiving the LA preparation had surface tension values of 24.6 mN·m−1 after adsorption, decreasing over 100 pulsations to values of 2.6±1.4 mN·m−1, significantly lower than the samples isolated from the animals given SA (p<0.05). A similar trend was observed for samples obtained from animals given the combinations of LA and SA. Analyses of the lavage fluid recovered from animals receiving radiolabelled SA revealed that 71.7±1.6% (15 mg·kg−1 SA group) and 74.0±2.9% (30 mg·kg−1 SA) of the 3H-radiolabel was recovered in the alveolar lavage.
Lung lavage analysis for total protein quantification based on a Lowry protein assay, and surface activity of recovered large aggregates (LA) (1 mg·mL−1) as assessed by the pulsating bubble surfactometer after 100 pulsations
Discussion
Alterations in the endogenous surfactant system have been shown to contribute to the lung dysfunction associated with ARDS 1, 16. Changes in the amount of surfactant aggregate forms within the airspace develop relatively early in the course of the injury, and progress as lung function deteriorates 3, 5. As a consequence, at the severe stages of lung injury, when patients exhibit decreased lung compliance, atelectasis and alveolar oedema, the surfactant isolated from these lungs is markedly dysfunctional, and consists predominantly of SA forms 4, 7, 17. It is unknown however, whether these increased SA pools are deleterious and contribute independently to lung dysfunction, possibly by interfering with the function of LA.
The in vitro functional assessments of the LA and SA utilized in the present study showed that SA had poor surface adsorption characteristics and surface tension-reducing activity, compared to LA which reduced surface tension to values near 0 mN·m−1. These results are consistent with previous findings 18. Interestingly, when SA were added to LA samples and functional activity was assessed, surface tension values were significantly different from both the LA or SA samples alone. Specifically, the function of the same concentration of LA that had reduced surface tension to values near 0 mN·m−1, was impaired when SA were added. These in vitro observations suggested that SA may interfere with the biophysical properties of LA, which the authors speculate occurred via mechanisms similar to surfactant inhibition via serum proteins as previously described in vitro 19.
Based on these results, the data obtained from the in vivo experiment was somewhat surprising, since animals receiving the combinations of LA and SA exhibited similar physiological responses as animals that received LA alone. Oxygenation values increased significantly following the instillation of these mixtures, while PIP values decreased. Given these results, and in comparison to the animals receiving SA alone, the authors conclude that at least in this experimental model, the presence of SA alone within the airspace had no effect on lung function in general, and did not interfere with the function of LA in vivo when administered together intratracheally.
When interpreting the results obtained from the in vivo and in vitro experiments in the present study, it is important to note the differences that exist between the two testing systems. Unlike the relatively controlled in vitro method used to analyse surface activity, the in vivo situation is dynamic and complex. In addition, the concentrations of surfactant aggregates tested in vitro may not reflect the specific proportions of aggregate forms existing within the alveolar subphase. The 15 LA/15 SA combination utilized in the current study was chosen to reflect the ratio of surfactant aggregate forms that is known to be present within the lung of a normal rat 14. The 15 LA/30 SA group represented the relative proportion of aggregate forms observed in animal models of severe ARDS 6. Once instilled into the lungs of these animals and assessed in vivo however, these aggregates may be distributed and oriented within the airspace in a considerably different manner to that which exists in the bubble surfactometer. Therefore, although the relative proportions of the aggregates were similar in the in vitro and in vivo experiments, the absolute concentrations of aggregate forms within the alveolar environment may have differed considerably, and consequently led to different results. Although the behaviour of surfactant at the alveolar level is complex, and beyond the scope of this study, the data involving the radiolabelled SA suggests that an increased uptake and/or clearance of SA from the airspace of these animals did not occur, as the majority of the 3H-labelled SA was recovered via lung lavage.
In vitro surface tension analyses of the LA obtained from the various groups following 60 min of ventilation were consistent with the in vivo observations. Specifically, LA isolated from animals given SA intratracheally had significantly higher surface tension values than LA isolated from animals given LA alone in vivo, but significantly lower than the SA preparations administered to these animals (p<0.001). This latter group of animals receiving LA had surface tension values comparable to samples obtained from groups receiving the combinations of LA and SA, consistent with the conclusion that SA had relatively little impact on the function of LA in vivo. In general, these results suggest that interactions of the administered aggregate forms with newly secreted endogenous surfactant, albeit the latter representing only a small quantity of material in this model, may have resulted in the surfactant “activation” phenomenon 20. Of note, it is unlikely that serum protein inhibition of surfactant within the airspace played a significant role in these observations as similar amounts of protein were recovered from all groups.
One important issue to consider when interpreting the current results is the methods used to purify the LA and SA preparations. LA were purified via sucrose gradient centrifugation and represented highly purified surfactant with its hydrophobic and hydrophilic components, and structural integrity maintained. Unfortunately, due to the large volume of the 40,000×g supernatant, the same methods could not be utilized to purify SA. Therefore, SA were concentrated using ultrafiltration. This method also co-purified large amounts of protein, and preliminary experiments showed that instillation of these SA into surfactant-deficient animals resulted in death after approximately 10 min of ventilation, presumably due to the overwhelming amount of protein present in this preparation (results not shown). To remove this protein from the SA, the preparation was organic solvent-extracted, which may also have changed the structure of the SA. Therefore, although the results of the present study represented the effects of the reconstituted hydrophobic components of the SA fraction, all methods currently used to isolate aggregate forms involve some degree of perturbation to the aggregate structure, and may not reflect the specific conformation of these aggregates in vivo.
In conclusion, this study supports the evidence that large aggregates are important for normal lung function. Furthermore, it was shown that although small aggregates interfered with the optimal function of large aggregates when tested in vitro, there were no obvious deleterious effects of increasing small aggregates quantities on large aggregates function in vivo. Further studies are required to explain the differences observed in the in vivo and in vitro results.
- Received February 1, 2001.
- Accepted July 25, 2001.
- © ERS Journals Ltd
References









