





Abstract
Inflammation and proteolytic processes play an important role in the progression of cystic fibrosis (CF) lung disease. The goal of this study was to describe bronchoalveolar lavage fluid (BALF) protein pattern of CF patients in comparison to controls and to assess if there is proteolytic degradation of surfactant protein A (SP-A), an important innate host defence component of the lungs.
BALFs from 17 clinically stable CF patients and from eight healthy children were separated by two-dimensional gel electrophoresis. Silver staining was used to show BALF proteins and Western blotting to detect SP-A isoforms.
In CF, BALF proteins of a low molecular weight ≤20 kD were more abundant than in controls. Various proteins were seen in CF which were not present in controls and vice versa. Degradation of SP-A was present in 15 of 17 CF BALFs but in none of the controls, in contrast polymeric isoforms were seen in all controls and in four of 17 CF patients.
Proteolytic damage to surfactant protein A and significant changes of normal bronchoalveolar lavage fluid proteins occur in lungs of cystic fibrosis patients. Identification of altered bronchoalveolar lavage fluid proteins may give new insights into pathogenic mechanisms and provide new targets for therapy.
This project was supported by a grant from the Wilhelm Sander Stiftung (Gr 93.002.1/2).
Cystic fibrosis (CF) is a lethal hereditary disorder caused by mutations of the CF transmembrane conductance regulator (CFTR) gene leading to abnormal exocrine gland secretion and chronic inflammation of the airways. A massive influx of activated neutrophils and high levels of proteolytic enzymes play a major role in the progression of CF lung disease and occur early in life 1–3. The airway of CF patients is susceptible to infection with various organisms, most notably Pseudomonas aeruginosa, Staphylococcus aureus and Haemophilis influenzae.
Bronchoalveolar lavage fluid (BALF) proteins are a potential source for detecting and characterizing biochemical alterations associated with airway diseases. In healthy subjects BALF proteins were predominantly identified as serum proteins 4, 5. Alterations of nasal fluid or BALF proteome (the set of proteins expressed in a specific compartment) have so far been reported for a limited number of respiratory diseases not including CF 5–8. In this study the differences between the BALF proteome of CF patients compared to controls were analysed. In addition the hydrophilic surfactant associated protein A (SP-A), which plays a central role in host defence in the air space 9 was focused on. In vivo studies confirmed the important role of SP-A in the innate immune system of the lung by demonstrating an increased susceptibility to P. aeruginosa and Group B Streptococci in SP-A knock out mice 10. Human SP-A is a glycoprotein composed of 18 polypeptide chains, each of about 34 kD molecular weight, isoelectric point 4.2–4.9. SP-A interacts with a variety of bacteria, viruses and fungi including CF associated pathogens as P. aeruginosa, S. aureus, H. influenza 9 and Aspergillus fumigatus 11 and enhances the uptake of some of these organisms by phagocytic cells 12. Adequate amounts of functional SP-A must be present in the lung to maintain host defence. In previous studies altered BALF concentrations of SP-A in CF have been demonstrated 13, 14. Based on in vitro studies 15, 16, it is likely that degradation of SP-A may occur in vivo in CF and this may contribute to impaired host defence in CF.
Alterations of the BALF proteome and of SP-A are presented in 17 CF patients compared to eight healthy children using high-resolution two-dimensional polyacrylamid gel electrophoresis (2D-PAGE).
Materials and methods
Patient population
BALF was obtained from eight healthy children and 17 patients with CF. The healthy subjects had no history of chronic respiratory disease or upper and lower tract infections in the 2-month period prior to participation in this study. These children were undergoing elective surgery for nonpulmonary diseases. After written informed consent, BAL was performed under general anaesthesia and tracheal intubation before surgery.
The 17 patients with CF were in a clinically stable condition and had no changes in therapy for >2 weeks. They were randomly selected from the CF outpatient department (table 1⇓). The diagnosis was established by typical clinical manifestation of the disease and confirmed by positive sweat tests in all CF patients. In the genetic analysis six of the 17 CF patients were homozygous for ΔF508, eight heterozygous and three showed other infrequent mutations. At the time of evaluation none of the subjects had an acute exacerbation of the lung disease. The BAL procedure was carried out under local anaesthesia. The study protocol was approved by the institutional review boards and written informed consent was obtained from the parents of all children and of older children before the study.
Clinical characteristics of cystic fibrosis patients and healthy controls
Bronchoalveolar lavage
BAL was performed in the right middle lobe with a flexible bronchoscope using 3 mL·kg−1 of 0.9% NaCl as previously described in detail 13. For further analysis BALF was filtered through one layer of loose, sterile gauze and centrifuged at 200×g for 10 min at 4°C immediately after BAL. To avoid ex vivo degradation of proteins, aliquots of BALF supernatant were immediately frozen at -70°C until assayed. The cell differential was evaluated using cytospin preparations of BALF and bacteriology was evaluated with standard techniques.
Determination of neutrophil elastase activity
Neutrophil elastase (NE) activity was determined spectrophotometrically using the specific substrate N-Suc-Ala-Ala-Ala-p-Nitroanilide (Elastin Products Co., Owensville, MO, USA) as described previously 13, 15. Assays were performed at room temperature on 96 well plates. The change of absorption was measured continuously for 5 min at 410 nm in an automated recording spectrophotometer (Anthos HT III, Anthos Labtech Instruments, Salzburg, Austria). NE activity was determined by comparing values obtained from the BALF samples to those of a concurrently run standard (875 U·mg−1, Elastin Products Co., Owensville, MO, USA). All samples were assessed in triplicate.
Two-dimensional electrophoresis
After the addition of Pefabloc protease inhibitor (Merck, Darmstadt, Germany), lavage fluid aliquots were desalted by extensive dialysis at 4°C (tubing cut-off 1 kD). Protein content was assessed before lyophilization and samples containing 80 µg of total protein were separated by horizontal high-resolution two-dimensional electrophoresis under reducing and denaturing conditions according to GÖrg 17. In preliminary experiments a high reproducibility for the location and abundance of the protein-spots on the gel was shown. Gels were silver stained 18 or proteins were transferred onto nitrocellulose membranes (Amersham Pharmacia Biotech, Uppsala, Sweden) for semi-dry Western blotting. SP-A was identified with a specific polyclonal antibody to human SP-A (1:5000; Byk Gulden, Konstanz, Germany). Purified human SP-A isolated from patients with pulmonary alveolar proteinosis served as a standard. Clara cell protein (rabbit anti-human urine protein 1), albumin and α1-protease inhibitor (α1-PI) were also identified with specific antibodies (Dako, Glostrup, Denmark). Enhanced chemiluminescence assay (Amersham Life Science, Amersham, Buckinghamshire, UK) with horseradish-peroxidase-conjugated goat antirabbit polyclonal immunoglobulin (Ig)-G (1:10,000; BioRad, Hercules, CA, USA) as a second antibody was used for detection.
Silver-stained gels and blots were scanned with a computing densitometer (Fluor-S MultiImager, Melanie 2.1 software (BioRad, Richmond, CA, USA)). To find corresponding protein-spots between gels, all silver-stained gels of each group (CF and control) were stacked and spots were matched according to molecular weight, isoelectric point standards and 15 hand-selected landmarks being present in each gel. Two “synthetic gels” were created with Melanie 2.1 software, one for CF and one for controls. These gels contained only the characteristic protein-spots for each group which have been detected in ≥85% of all gels in each group 19. These synthetic gels were matched to gels of the other group (e.g. CF versus control) to identify differences in protein-spot patterns. Direct quantification of proteins in silver-stained gels or Western blot analysis were performed by measuring the relative volume (Vol%) of the spots. The Vol% was calculated by dividing the Vol of a single spot through the sum of the Vol of all spots of a gel times 100. The Vol of a spot is defined as ∑ I (x,y), where I (x,y) is defined as the intensity of a point p (x,y) being part of the spot. I (x,y) is calculated by the software dependent on the saturation of the spot with respect to silver staining properties 19.
Statistical analysis
Results are expressed as the mean±sem for n independent determinations. Comparisons for continuous variables were made by an unpaired two-tailed t-test or for frequencies by the Chi-squared test, correlation analysis was performed by calculating the Pearson correlation coefficient.
Results
The total and differential cell counts of the BALF in CF differed significantly from control BALF (table 1⇑) showing a well-known neutrophil-dominated inflammation 1, 3. In the cell free BALF supernatant free NE was present in 14 of 17 CF patients but in none of the controls (CF 10.4±2.8 U·mL−1). The elastase activity was correlated with the neutrophil cell count in BALFs (r=0.6; p=0.014). Total protein content was elevated in BALF of CF compared to controls (CF 314±46 µg·mL−1; control 105±32 µg·mL−1; p=0.007) and was correlated with the total cell count (r=0.75; p=0.0006), the neutrophil cell count (r=0.77; p=0.0006), and the NE activity in BALF (r=0.71; p=0.0014).
Bronchoalveolar lavage proteome
The BALF proteins of all CF patients and controls were separated by 2D-PAGE and silver-stained. The protein display of CF BALF showed major differences from that of controls (table 2⇓, fig. 1⇓). In control subjects albumin was the most abundant single protein (26.3±3% as estimated in Vol% of total silver-stained proteins) in BALF. Western blot analysis with antibodies against human serum albumin showed, that albumin was infrequently present in CF BALF. No proteolytic fragments of albumin were found. In contrast in CF BALF α1-PI was the most predominant protein (9.3±1.9 Vol%) but was uncommonly seen in controls (table 2⇓, fig. 1⇓). The Vol% of α1-PI has been found to correlate with the activity of NE (U·mL−1) in BALF (r=0.75; p<0.0005). Low molecular weight (LMW) proteins were more abundant on silver-stained gels in CF than in controls (fig. 1⇓). Therefore, two groups of proteins with respect to high (>20 kD) or low (≤20 kD) molecular weight were arbitrarily created in each gel. In CF BALF, LMW protein-spots of ≤20 kD represented >40% of visible spots and nearly 50% of total protein (as estimated in Vol% of total silver-stained proteins) being 3–7-fold elevated compared to control BALFs (table 2⇓; fig. 1⇓). There was a positive correlation between the Vol% of LMW proteins ≤20 kD and the neutrophil cell count in BALF (r=0.6; p=0.015). There was no significant correlation noted to NE activity, forced expiratory volume in one second (FEV1), the age at BAL or total protein content. CF patients showing a positive bacterial culture of BALF (n=10) had significantly more spots in the LMW ≤20 kD than those without bacteriological findings (43.2± 2.1% versus 35.3±2.8% of total number of spots on gel; p=0.025).

Representative high-resolution two-dimensional polyacrylamid gel electrophoresis (2D-PAGE) pattern of bronchoalveolar lavage fluid (BALF) proteins of a) a healthy child and b) a cystic fibrosis (CF) patient showing moderate human neutrophil elastase activity (3.9 U·mL−1). Separated proteins were detected by silver staining. The 2 D-PAGE was performed under reducing conditions on a linear immobilized pH gradient pH 3–10, followed by a 12% SDS-PAGE. The figure contains scanned gels, presented as an unmanipulated image, except for overall contrast using programs for graphic processing. Identified proteins are indicated with numbers. 1: surfactant protein A*; 2: albumin*; 3: immunglobulin A α chain*; 4: immunglobulin G heavy chain*; 5: immunglobulin G light chains κ,λ*; 6: α1-PI+. Proteins were identified by *: comparison with a master plasma pattern in Swiss-2D PAGE database; +: western immunoblotting.
Analysis of silver-stained proteins separated by high-resolution two-dimensional polyacrylamid gel electrophoresis (2D-PAGE)
Some proteins were seen in CF BALF, but not in those of controls and vice versa. To localize these proteins on the gels the Melanie 2.1 software package (Biorad, Richmond, CA, USA) was used 19. Two synthetic gels were created (fig. 2⇓), one for controls containing all spots being present in ≥7 of 8 control gels (67 spots) and one for CF, containing those protein spots being present in ≥15 of 17 of CF gels (76 spots). Afterwards, these two resulting synthetic gels were matched to all native silver-stained gels of the other group. Nineteen protein spots were unique for the CF group, 14 of these being located in the LMW ≤20 kD (fig. 2⇓). In contrast, 14 protein spots of the high molecular weight range were present in ≥7 of 8 of the control gels, but in none of CF gels (fig. 2⇓).

Two synthetic 2D-gels created by Melanie 2.1 software containing only characteristic protein spots for each group: a) cystic fibrosis (CF) and b) control. Scanned, silver-stained gels of each bronchoalveolar lavage fluid (BALF) were analysed by the software and spots being present in ≥7 of 8 control gels were copied in the control synthetic gel, spots being present in ≥15 of 17 of CF gel were copied in the CF synthetic gel. Each of the two resulting synthetic gels were matched to all silver-stained gels of the other group to find protein spots being characteristic for the one group and not being present in the other group. Unique characteristic protein spot, for which no corresponding protein spot can be found in gels of the other group, indicated by arrows. 1: surfactant protein A; 2: albumin ; 3: immunglobulin A α chain ; 4: immunglobulin G heavy chain; 5: immunglobulin G light chains κ,λ ; 6: α1-PI.
Analysis of surfactant protein A
Although the amount of SP-A in BALF did not vary significantly between CF and controls, remarkable qualitative changes for SP-A were found. Enriched SP-A, which is associated with the 40,000 g BALF pellet was separated by 2D-PAGE under reducing conditions on immobilized pH gradient (IPG) strips pH 4–5.5. Silver staining of these gels showed a characteristic chain of 16–18 single protein-spots with a molecular weight of 31–35 kD and isoelectric points of 4.2–4.9 in both the CF (n=3) and control (n=2) groups (fig. 3⇓). In CF additional protein-spots with a molecular weight of ∼14–24 kD and pH of 4.5–5.1 were noted. 2D-PAGE gels of the same samples of BALF run in parallel were subjected to Western blotting and it was demonstrated, that these LMW protein-spots were immunoreactive with a polyclonal antibody to SP-A, as were the higher MW protein spots. Systematic Western blot analysis of all native BALF supernatants on IPG pH 3–10 demonstrated that LMW degradation products were present in 15 of 17 CF patients in varying quantity, but in none of the eight controls (table 3⇓, fig. 4⇓). In contrast, polymeric forms of SP-A were observed in all controls but in only four of 17 CF patients. These polymers of SP-A comprised of a molecular weight from ∼60 kD up to >100 kD and could not be reduced despite addition of increasing amounts of reducing agents (data not shown). The Vol% of SP-A degradation products correlated to the number of total cells in BALF (r=0.5; p=0.045), but not significantly to NE activity or neutrophil cell count. CF patients showing a positive bacterial culture of BALF (n=10) had more degradation of SP-A than those without bacteriological findings, not reaching significance (16.2±4.3 versus 7.4± 2.3 Vol% of total SP-A; p=0.13).

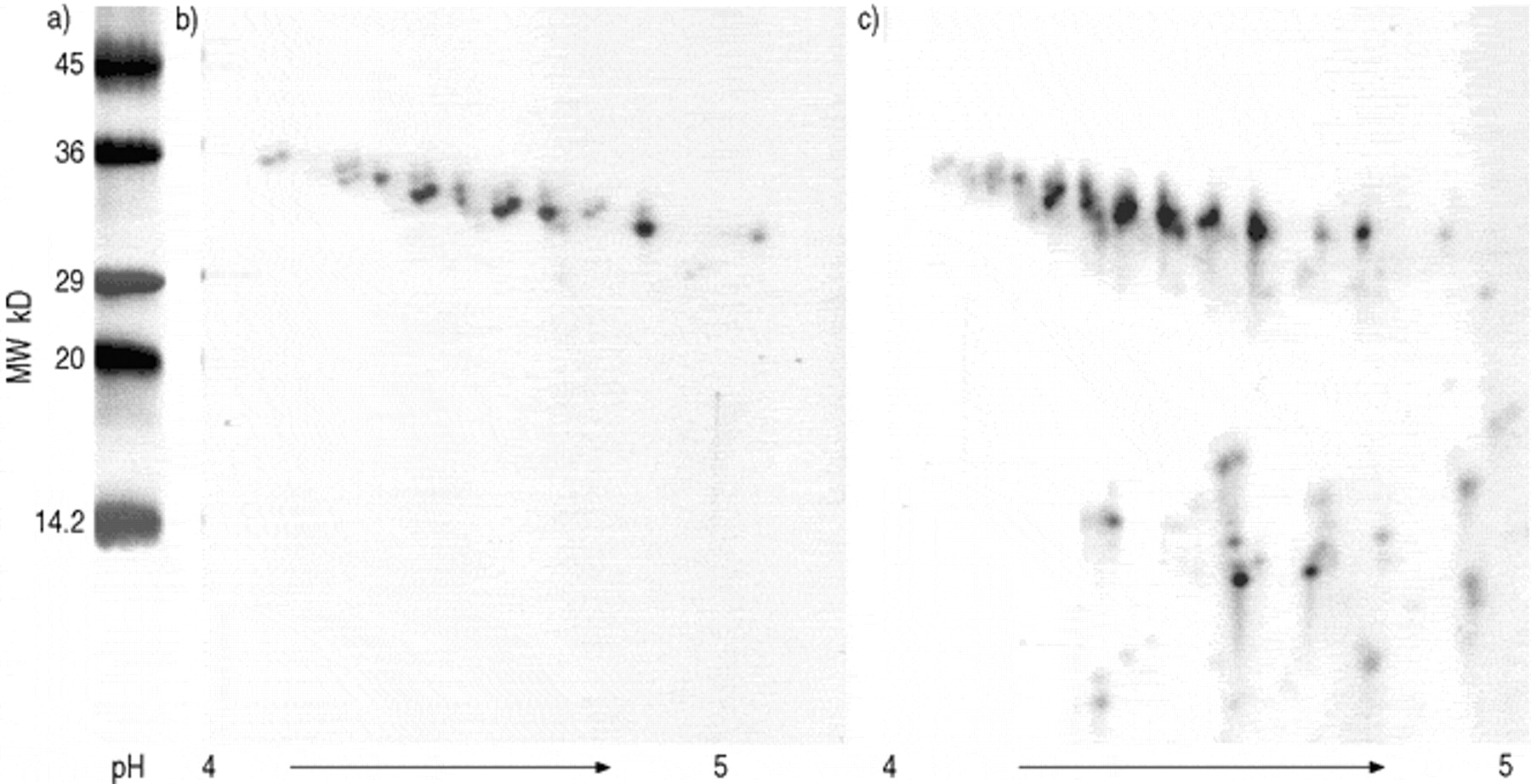
a) Molecular weight standard; b) surfactant protein A (SP-A) of a healthy child; c) SP-A of a cystic fibrosis (CF) patient. SP-A associated to the pellet of bronchoalveolar lavage fluid (BALF) after 40,000×g centrifugation was separated under reducing conditions by high-resolution two-dimensional polyacrylamide gel electrophoresis (2D-PAGE) (linear immobolized pH gradient pH 4–5.5) and detected by silver staining. The presence of SP-A was confirmed by Western blotting with polyclonal antibodies raised against human SP-A on an equal gel run in parallel.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Immunochemical staining patterns of surfactant protein A (SP-A) in bronchoalveolar lavage fluid (BALF) separated by (2D-PAGE), immobolized pH gradient pH 3–10 linear. Nitrocellulose blots were probed with polyclonal antibodies raised against human SP-A, chemiluminescence was used for detection. a) One dimensional separated isolated human SP-A as a standard; b) two dimensional separated SP-A of BALF proteins from two representative control subjects; c) two dimensional separated SP-A from four representative patients with CF showing different human leukocyte elastase (HLE) activities in BALF; I: patient with 0.9 U·mL−1 HLE activity; II: patient with 3.5 U·mL−1 HLE activity; III: patient with 3.9 U·mL−1 HLE activity; IV: patient with 18.6 U·mL−1 HLE activity; a: Di-and polymeric forms of SP-A, MW ~66–100 kD; b: monomeric form of SP-A ~30–36 kD, pI 4.2–4.9; c: degradation products of SP-A MW ~14–24 kD.
Analysis of surfactant protein A (SP-A) by high-resolution two-dimensional polyacrylamid gel electrophoresis
Discussion
In this study it has been shown, that significant differences in the overall BALF protein-patterns and qualitative alterations of SP-A in CF adolescents with clinical stable disease compared to healthy controls exist. Besides an elevated content of total protein, the BALF protein-pattern of CF patients differed significantly to that of controls, the latter showing a well-known concordance to the proteome of human serum 4–7.
In CF BAL and sputum a variety of proteases are elevated and are likely to play a role in the progression of the lung disease. Among these NE is one of the most abundant active neutrophil proteases. Thus in this study the findings were correlated to the activity of NE in BALF as a common representative of neutrophil derived proteolytic enzymes.
To better describe the proteome alterations in BALF, two different groups of proteins according to low (≤20 kD) and high (>20 kD) molecular weight were arbitrarily created. The predominance of the LMW (≤20 kD) proteins in CF BALF may at least partly be due to an accumulation of degraded structure, defence, bacterial or inflammatory proteins. This is in accordance with the data of a previous report by the authors, where an 8 week inhalation of α1-PI in CF patients led to a significant reduction in the number and Vol% of these LMW proteins ≤20 kD 20. Various proteins were seen in CF, which were not present in controls and vice versa. These proteins need to be identified in future studies as they allow a quantification of the relative abundance of various components involved in the inflammatory and proteolytic processes in CF lung disease. The primary goal of this study was to establish the overall protein pattern in CF and to localize potential targets for sequencing and protein identification.
In accordance with previous reports of elevated α1-PI levels in CF sputum the Vol% of α1-PI of total protein was much higher in CF BALF than in controls 3, 21. As the Vol% of α1-PI was strongly correlated to the NE activity in BALF, α1-PI must have been inactivated by proteolysis or oxidation 3, 21. The latter clearly predominated, as no degradation products of α1-PI were found by Western blot analysis (data not shown).
In addition to the altered protein-pattern of CF BALF qualitative changes for SP-A were found. SP-A was degraded to LMW products in the majority of BALFs of CF patients. Higher molecular forms were not present in CF BALF but were in all the controls. High resolution 2D-PAGE was used to describe in detail the human SP-A isomers of BALF of control and CF patients. The molecular weight and pI of the main chain of SP-A isomers are not altered in CF and are also in accordance with reports in the literature 22, 23. Baker et al. 24 and Doyle et al. 25 previously described the qualitative impairment of SP-A in BALF of patients with acute respiratory distress syndrome 25 and of patients with pulmonary alveolar proteinosis 25 compared to controls. Both found alterations in the distribution between polymeric and isomeric forms of SP-A, but no LMW products of SP-A (MW <24 kD). As confirmed by Western blotting, these proteolytic fragments of SP-A with a molecular weight of ∼14–24 kD were present in the majority of CF patients but in none of the controls. As there is a correlation of Vol% SP-A degradation products to the total number of cells in BALF, SP-A is likely to be damaged by cell-related proteolytic enzymes as NE or other proteases as Cathepsin G or B and collagenases. In vitro studies suggest, that rat and dog SP-A can be cleaved by NE 13, 14 and collagenase 26 resulting in proteolytic fragments of 21 kD (NE) or 20–23 kD and pI of 4.2–4.8 (collagenase). In these studies the biophysical function of surfactant in surface adsorption was impaired after exposure to the above proteases. Thus it is likely that the impaired biophysical function of surfactant in CF may in part be related to proteolytic damaged SP-A. Loss of the host defence properties with impaired SP-A is likely, but has to be demonstrated in future studies.
The high molecular assemblies (>60 kD) of SP-A were found in all BALFs from controls but almost not in CF. These forms of SP-A were likely to represent dimers and oligomers 25, but it is not known how they are generated and what their exact functional role is. A quantitative reduction of SP-A in BALF from CF patients with severe lung disease has been reported previously 13, 14, whereas in asymptomatic CF infants no alterations and with infection an upregulation of SP-A has been noted 27. Reduction of SP-A with advanced disease may be due to a reduced synthesis or excessive proteolytic degradation of SP-A. In the latter case the generated fragments must be small enough not to be detected by specific antibodies in enzyme linked immunosorbent assay (ELISA). The proteolytic fragments of 14–24 kD observed in this study will still be reactive in ELISA tests using the same antibodies as in Western blot. Therefore, the total amount of intact SP-A will be overestimated in ELISA.
Interestingly elevated LMW proteins, SP-A fragments and NE activity have been observed in the absence of bacterial or fungal findings in CF BALF. This indicates that inflammation and subsequent damage of lung proteins can occur independent of bacterial findings in CF lung.
It is concluded, that proteolytic damage of surfactant protein-A and the changes of other bronchoalveolar lavage fluid proteins are likely to be due to a neutrophil dominated inflammation and might contribute to a reduced ability in cystic fibrosis to remove the specific pathogens from their lungs. After the identification and characterization of the proteins expressed under various clinical conditions, therapeutic interventions aimed at reducing the inflammation and the proteolytic activity in the airspace may be monitored by the changes in the proteome display.
- Received August 30, 2000.
- Accepted December 8, 2000.
- © ERS Journals Ltd
References









