





Abstract
Chronic obstructive pulmonary disease (COPD) is the third leading cause of morbidity and death globally. The lack of effective treatments results from an incomplete understanding of the underlying mechanisms driving COPD pathogenesis.
Interleukin (IL)-22 has been implicated in airway inflammation and is increased in COPD patients. However, its roles in the pathogenesis of COPD is poorly understood. Here, we investigated the role of IL-22 in human COPD and in cigarette smoke (CS)-induced experimental COPD.
IL-22 and IL-22 receptor mRNA expression and protein levels were increased in COPD patients compared to healthy smoking or non-smoking controls. IL-22 and IL-22 receptor levels were increased in the lungs of mice with experimental COPD compared to controls and the cellular source of IL-22 included CD4+ T-helper cells, γδ T-cells, natural killer T-cells and group 3 innate lymphoid cells. CS-induced pulmonary neutrophils were reduced in IL-22-deficient (Il22−/−) mice. CS-induced airway remodelling and emphysema-like alveolar enlargement did not occur in Il22−/− mice. Il22−/− mice had improved lung function in terms of airway resistance, total lung capacity, inspiratory capacity, forced vital capacity and compliance.
These data highlight important roles for IL-22 and its receptors in human COPD and CS-induced experimental COPD.
Abstract
IL-22 and its receptors are increased in both human and experimental chronic obstructive pulmonary disease (COPD). IL-22 drives neutrophilic inflammation and impaired lung function in experimental COPD. http://bit.ly/2Vsri6T
Introduction
Chronic obstructive pulmonary disease (COPD) is the third leading cause of morbidity and death and imposes a significant socioeconomic burden globally [1]. It is a complex, heterogeneous disease characterised by chronic pulmonary inflammation, airway remodelling and emphysema, which are associated with progressive lung function decline [2]. Cigarette smoke (CS) is a major risk factor for COPD [2]. The mainstays of therapy for COPD are glucocorticoids, β2-adrenergic receptor agonists and long-acting muscarinic antagonists [3]. However, these agents only provide symptomatic relief rather than modifying the causal factors or suppressing disease progression [3]. There is emerging interest in altered lung and gut microbiomes and the gut–lung axis which could be modified for therapeutic gain [4, 5]. However, there is currently a lack of effective treatments for COPD due to the poor understanding of the underlying mechanisms.
Interleukin (IL)-22 is a member of the IL-10 cytokine family which is implicated in several human diseases, including mucosal-associated infections and inflammatory disorders of the lung [6]. CD4+ T-helper cells, γδ T-cells, natural killer (NK)T-cells and group 3 innate lymphoid cells (ILC3) are generally the major cellular sources of IL-22 [6]. Unlike IL-22, expression of the IL-22 receptor (IL-22R) is largely restricted to structural cells. This ligand–receptor distribution permits immune cells to regulate responses of stromal cells, and particularly at barrier surfaces such as the lung, where epithelial cells play an active role in initiating, regulating and resolving immune responses. IL-22R is a cell-surface heterodimer consisting of IL-22RA1 and IL-10RB [6]. IL-22RA2 is a naturally occurring IL-22 antagonist which negatively regulates IL-22-induced inflammatory responses [6, 7]. Functional studies in murine systems indicate that IL-22 has immune-regulatory properties in infection, inflammation, autoimmunity and cancer [6]. In these models, the functional consequences of IL-22 expression can be either pathological or protective, depending on the context in which it is expressed. Indeed, increased IL-22 levels and IL-22+ cells have been demonstrated in the blood, sputum and lung biopsies of COPD patients [8]. The role of IL-22 in lung antimicrobial defence and the impact of COPD on this defence pathway has been reported [9, 10]. In experimental COPD, Haemophilus influenzae infection impaired IL-22 production, and wild-type (WT) and IL-22-deficient (−/−) mice had impaired clearance [10]. CS exposure suppressed Streptococcus pneumoniae-induced IL-22 production and treatment with recombinant IL-22 restored bacterial clearance [11]. Despite this, there is limited knowledge of the role that IL-22 plays in COPD pathogenesis independent of respiratory infection.
Here, we investigate its role using gene expression analysis of airway epithelial brushings and parenchymal cores from human COPD patients, an established mouse model of CS-induced experimental COPD that recapitulates the critical features of human disease [4, 12–18], and IL-22 reporter and Il22−/− mice [19]. IL-22 and IL-22R mRNA and protein were increased in the airways of mild-to-moderate COPD patients. IL-22 and IL-22+ T-cells and ILC3s were increased in experimental COPD. CS-induced pulmonary neutrophilic inflammation, airway remodelling and emphysema were reduced and lung function was improved in Il22−/− mice compared to WT controls, thus implicating IL-22 in COPD pathogenesis.
Methods
Ethics statement, animal details, additional methods and statistical analyses are described in the supplementary material.
Human gene expression
Analysis of IL22, IL22RA1, IL10RB and IL22RA2 in published human array datasets (accession numbers: GSE5058 and GSE27597) [20–22] was performed using Array Studio software (OmicSoft Corporation, Research Triangle Park, NC, USA).
Mice
Animals were female, 7–8-week-old, WT C57BL/6 mice, Il17aeGFP/+;Il22td−tomato/+ reporter and Il22−/− mice on a C57BL/6 background [19].
Experimental COPD
Mice were exposed to normal air or nose-only inhalation of CS for 8 weeks in a protocol representative of a pack-a-day smoker as extensively described previously [4, 12–18, 23, 24].
Quantitative PCR
Total RNA was extracted from whole lung tissue and blunt-dissected airways and parenchyma and reverse transcribed [13]. mRNA transcripts were determined using real-time quantitative (q)PCR (ABIPrism7000, Applied Biosystems, Scoresby, Victoria, Australia) using custom-designed primers (Integrated DNA Technologies, Baulkham Hills, New South Wales, Australia) (supplementary table S1).
Flow cytometry
IL-17A+ and IL-22+ CD4+ T-cells, γδ T-cells, NKT-cells and ILC3s in lung homogenates were determined based on surface marker expression (supplementary table S2) [25–27] using a BD FACSAriaIII. Flow cytometry antibodies were from Biolegend (Karrinyup, Australia) or BD Biosciences (North Ryde, Australia) (supplementary table S3, supplementary figure S1).
Pulmonary inflammation
Airway inflammation was assessed by differential enumeration of inflammatory cells in bronchoalveolar lavage fluid (BALF) [12, 14, 28, 29]. BALF supernatants were stored at −20°C for assessment of IL-22 protein levels. Tissue inflammation was assessed by enumeration of inflammatory cells [12–14, 29] and histopathological scoring based on established criteria [30].
ELISA
IL-17A, IL-22, myeloperoxidase (MPO) and neutrophil elastase protein levels were quantified using commercially available ELISA kits (R&D Systems or Biolegend) [19].
Immunohistochemistry
Lungs were perfused, inflated, formalin fixed, paraffin embedded and sectioned (4 μm) [13, 14]. Longitudinal sections of the left lung were deparaffinised and stained with antibodies against IL-22RA1 or IL-22RA2. Immunohistochemistry (IHC) in human samples is described in supplementary tables S4–S6 [31].
Airway remodelling
Airway epithelial (μm2) and collagen deposition area (μm2) were assessed in a minimum of four small airways (basement membrane perimeter <1000 μm) per section [12–14, 17, 18]. Data were quantified using ImageJ software (version 1.50; National Institutes of Health, Bethesda, MD, USA) and normalised to basement membrane perimeter (μm).
Alveolar enlargement
Alveolar diameter was assessed using the mean linear intercept technique [12–14, 17, 18, 32].
Lung function
Mice were anaesthetised using ketamine (100 mg·kg−1) and xylazine (10 mg·kg−1), tracheas cannulated and attached to Buxco® Forced Maneuvres apparatus (DSI, St. Paul, MN, USA) to assess total lung capacity (TLC) [12, 13]. FlexiVent apparatus (FX1 System; SCIREQ, Montreal, Canada) was used to assess lung volume, airway resistance, inspiratory capacity, forced vital capacity (FVC), compliance and elastance (tidal volume 8 mL·kg−1, respiratory frequency 450 breaths·min−1) [12, 33, 34].
Results
IL-22 and IL-22R mRNA expression and protein levels are increased in human COPD
First, we determined whether the mRNA expression of IL-22 and its receptors IL-22RA1 and IL-10RB and antagonist IL-22RA2 were altered in humans with mild-to-moderate COPD (Global Initiative for Chronic Obstructive Lung Disease (GOLD) stage I or II accession GSE5058 [20, 21, 35]). Pre-existing microarray data from airway epithelial brushings of healthy nonsmokers, healthy smokers and COPD patients were interrogated [20]. IL-22, IL-22RA1, IL-10RB and IL-22RA2 mRNA expression were not significantly altered in airway epithelial brushings from healthy smokers compared to nonsmokers (figure 1a–d). However, importantly, IL-22 (2.01-fold), IL-22RA1 (2.48-fold), IL-10RB (3.26-fold) and IL-22RA2 (1.78-fold) mRNA expression was increased in airway epithelial brushings from patients with mild-to-moderate COPD compared to nonsmokers. Similar results were observed when mild-to-moderate COPD was compared to healthy smokers.

Interleukin (IL)-22 and IL-22R mRNA expression are increased in airway epithelial brushings from human mild-to-moderate chronic obstructive pulmonary disease (COPD) patients compared to healthy smokers and nonsmokers. Microarray data from airway epithelial cells from healthy human nonsmokers, healthy smokers without COPD and COPD patients with Global Initiative for Chronic Obstructive Lung Disease (GOLD) stage I (mild) or II (moderate) disease (accession: GSE5058 [20]) were interrogated: a) IL-22, b) IL-22RA1, c) IL-10RB, d) IL-22RA2 mRNA expression. Microarray data from lung parenchymal cores from human healthy nonsmokers and COPD patients with GOLD stage IV (severe) disease (accession: GSE27597 [22]) were interrogated: e) IL-22, f) IL-22RA1, g) IL-10RB, h) IL-22RA2 mRNA expression. Data are expressed as log2 intensity robust multi-array average signals. The Benjamini–Hochberg method for adjusted p-value/false discovery rate was used to analyse differences between nonsmokers, smokers and COPD patients. RMA: robust multichip averaging; ns: nonsignificant. #: p<0.005 compared to COPD.
We then assessed the mRNA expression of IL-22 and its receptors in pre-existing microarray data from lung parenchyma cores from severe COPD patients (GOLD stage IV [35] accession: GSE27597 [22]). There was no change in IL-22, IL-22RA1, IL-10RB or IL-22RA2 expression in cores from COPD patients compared to nonsmokers without COPD (figure 1e–h). IL-22, IL-22RA1, IL-22RA2 and IL-10RB were unchanged in peripheral lung tissue from patients with mild emphysema (supplementary figure S2 from GSE8581). There was no significant correlation between pack-years and IL-22, IL-22RA1 and IL-22RA2 gene expression in lung tissue (supplementary figure S3 from GSE17770). Using lung cancer as a disease control, no differential expression of IL-22, IL-22RA1, IL-22RA2 or IL-10RB in either bronchial brushings (GSE4115) or lung tissue (GSE1650) between healthy smokers and subjects with lung cancer were observed (supplementary figures S4 and S5).
Finally, we assessed IL-22 and receptor protein levels in human COPD using IHC. The percentage of IL-22+ alveolar macrophages and IL-22RA1+ and IL-10RB+ airway epithelial cells were increased in COPD compared to age- and smoking history-matched smokers with normal lung function (figure 2, supplementary figure S6 and supplementary tables S6–S9). No change in IL-22RA2 was detected (supplementary table S8).
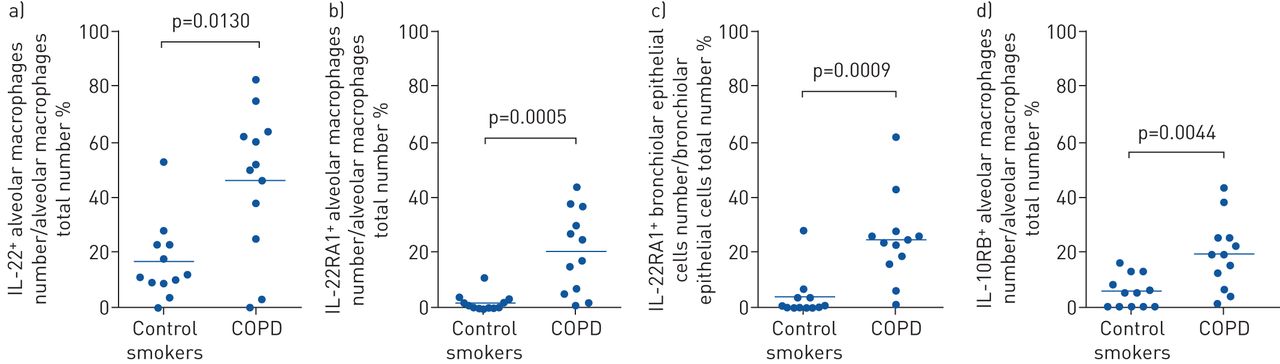
Interleukin (IL)-22, IL-22RA1 and IL-10RB, but not IL-22RA2 protein are increased in human chronic obstructive pulmonary disease (COPD). IHC for IL-22 and its receptors in peripheral lung from smokers with mild-to-moderate stable COPD and compared to age- and smoke history-matched smokers with normal lung function. a) IL-22+ alveolar macrophages; b) IL-22 receptor (IL-22R)A1+ alveolar macrophages; c) IL-22RA1+ airway epithelial cells; d) IL-10RB+ alveolar macrophages. Data are presented as mean±sem, n=12 per group.
In a separate cohort of COPD patients, IL-22RA1 was increased in airway epithelial cells of current smokers with COPD compared to nonsmokers (supplementary figure S7 and supplementary table S10). When combined with ex-smokers with COPD, this IL-22RA1 signal in the airway epithelium is lost (supplementary figure S7).
IL-22 and receptor protein levels are increased in the lungs in experimental COPD
Next, we investigated the expression of IL-22 and its receptors in CS-induced experimental COPD, which models mild-to-moderate COPD. We first confirmed that IL-22 was increased in experimental COPD. Il22 mRNA was difficult to detect in mouse lungs; therefore, we assessed protein levels using ELISA in both whole-lung homogenates (include both airways and parenchyma) and BALF supernatants. CS-exposure of WT mice resulted in increased IL-22 protein levels in lung homogenates, but not BALF supernatants compared to normal air-exposed controls (figure 3a and b). IL-22 protein levels were unaltered following 1 week of CS exposure (supplementary figure S8). Collectively, these data show that IL-22 is increased in both human and experimental COPD and are consistent with previous reports [8].

Interleukin (IL)-22 protein levels are increased in the lungs of cigarette smoke (CS)-exposed mice with experimental chronic obstructive pulmonary disease. Wild-type (WT) C57BL/6 mice were exposed to normal air or CS for 8 weeks. IL-22 protein levels in a) lung homogenates and b) bronchoalveolar lavage fluid supernatants were assessed using ELISA. In separate experiments, airways and parenchyma were blunt-dissected and IL-22 receptor mRNA expression assessed. Airway c) Il22ra1, d) Il10rb and e) Il22ra2 and parenchymal f) Il22ra1, g) Il10rb and h) Il22ra2 mRNA expression. Data are presented as mean±sem, n=6, with another independent experiment showing similar results. Two-tailed Mann–Whitney t-test was used to analyse differences between two groups. *: p<0.05 compared to normal air-exposed WT controls; ns: nonsignificant.
Next, we assessed IL-22 receptor expression in blunt-dissected airways versus parenchymal tissue [13]. CS-exposure had no statistically significant effect on Il22ra1 or Il10rb mRNA expression, but did reduce Il22ra2 expression in the airways compared to normal air-exposed controls (figure 3c–e). CS exposure also did not affect Il22ra1 or Il22ra2 mRNA expression, but did increase Il10rb expression in the parenchyma compared to normal air-exposed controls (figure 3f–h). While no statistically significant differences in Il22ra1 mRNA expression were observed in this model, it is notable that Il22ra1 mRNA expression was ∼10-fold higher in the airways than the parenchyma.
Finally, we assessed IL-22 receptor protein expression in mouse lung tissue sections. CS-exposure resulted in notable increases in both IL-22RA1 and IL-22RA2 protein levels, particularly in airway epithelial cells, but also in alveolar macrophages (supplementary figure S9).
IL-22+ CD4+ T-cells, γδ T-cells, NKT-cells and ILC3s are increased in the lungs in experimental COPD
Given that IL-22 is increased in both human and experimental COPD, we defined the cellular source of increased pulmonary IL-22 using Il17aeGFP/+; Il22td-tomato/+ reporter mice which enable the detection of IL-17A+ and IL-22+ cells without ex vivo stimulation. CS-exposure of reporter mice resulted in increased numbers of IL-17A+, IL-22+ and IL-17A+IL-22+ CD4+ T-cells, γδ T-cells, NKT-cells and ILC3s compared to normal air-exposed controls (figure 4a–p). We then assessed the relative proportions of these cells following CS-exposure (figure 4q–s). As shown previously [36], γδ T-cells were the dominant source of IL-17A following CS exposure (figure 4q). CD4+ T-cells, NKT-cells and ILC3s were the major IL-22-producing cells (figure 4r), while NKT-cells were the dominant source of dual IL-17A+IL-22+ cells (figure 4s).

(previous page) Interleukin (IL)-22+ CD4+ T-cells, γδ T-cells, natural killer (NK)T-cells and group 3 innate lymphoid cells (ILC3s) are increased in the lungs of cigarette smoke (CS)-exposed mice with experimental chronic obstructive pulmonary disease. Il17aeGFP/+;Il22td−tomato/+ reporter mice were exposed to normal air or CS for 8 weeks and the cellular source of IL-17A and IL-22 in the lung was assessed using flow cytometry. Total numbers of a) IL-17A+, b) IL-22+ and c) IL-17A+IL-22+ CD4+ T-cells in the lung. Total numbers of d) IL-17A+, e) IL-22+ and f) IL-17A+IL-22+ γδ T-cells in the lung. Total numbers of g) IL-17A+, h) IL-22+ and i) IL-17A+IL-22+ NKT-cells in the lung. Total numbers of j) IL-17A+, k) IL-22+ and l) IL-17A+IL-22+ ILC3 cells in the lung. Relative proportions of CD4+ T-cells, γδ T-cells, NKT-cells and ILC3s expressing m) IL-17A, n) IL-22 and o) IL-17 and IL-22. Data are presented as mean±sem, n=6, with another independent experiment showing similar results. Two-tailed Mann–Whitney t-test was used to analyse differences between two groups. *: p<0.05 compared to normal air-exposed controls. Representative fluorescence-activated cell sorting (FACS) plots of IL-17A+ and IL-22+ CD4+ T-cells; IL-17A+ and IL-22+ γδ T-cells; IL-17A+ and IL-22+ NKT-cells; and IL-17A+ and IL-22+ ILC3s are presented in supplementary figure S10.
CS-induced pulmonary neutrophils were reduced in Il22−/− mice
We next investigated whether IL-22 plays a role in the pathogenesis of experimental COPD. WT and Il22−/− mice were exposed to normal air or CS for 8 weeks [12–18]. Pulmonary inflammation in BALF was assessed by staining and differential enumeration of inflammatory cells. CS exposure of WT mice resulted in significantly increased total leukocytes, macrophages, neutrophils and lymphocytes compared to normal air-exposed WT controls (figure 5a–d). CS-exposed Il22−/− mice also had increased numbers of these cells compared to normal air-exposed Il22−/− controls. Neutrophils were significantly reduced, but total leukocytes, macrophages and lymphocytes were unaltered in CS-exposed Il22−/− mice compared to CS-exposed WT controls.

Cigarette smoke (CS)-induced pulmonary inflammation is reduced in Il22−/− mice. Wild-type (WT) and interleukin (IL)-22-deficient (Il22−/−) C57BL/6 mice were exposed to normal air or CS for 8 weeks to induce experimental chronic obstructive pulmonary disease. a) Total leukocytes, b) macrophages, c) neutrophils and d) lymphocytes in bronchoalveolar lavage fluid (BALF); e) representative images of parenchymal inflammatory cells; f) numbers of parenchymal inflammatory cells per high powered field; g) representative images of lung histopathology scoring; h) total histopathology score in lung sections and scores specifically in the i) airway, j) vascular and k) parenchymal regions; l) Cxcl1, m) Cxcl2 and n) Il17a mRNA expression in lung homogenates. Data are presented as mean±sem, n=6, with another independent experiment showing similar results. The one-way ANOVA with Bonferroni post-test analysed differences between three or more groups. *: p<0.05 compared to normal air-exposed controls; ns: nonsignificant.
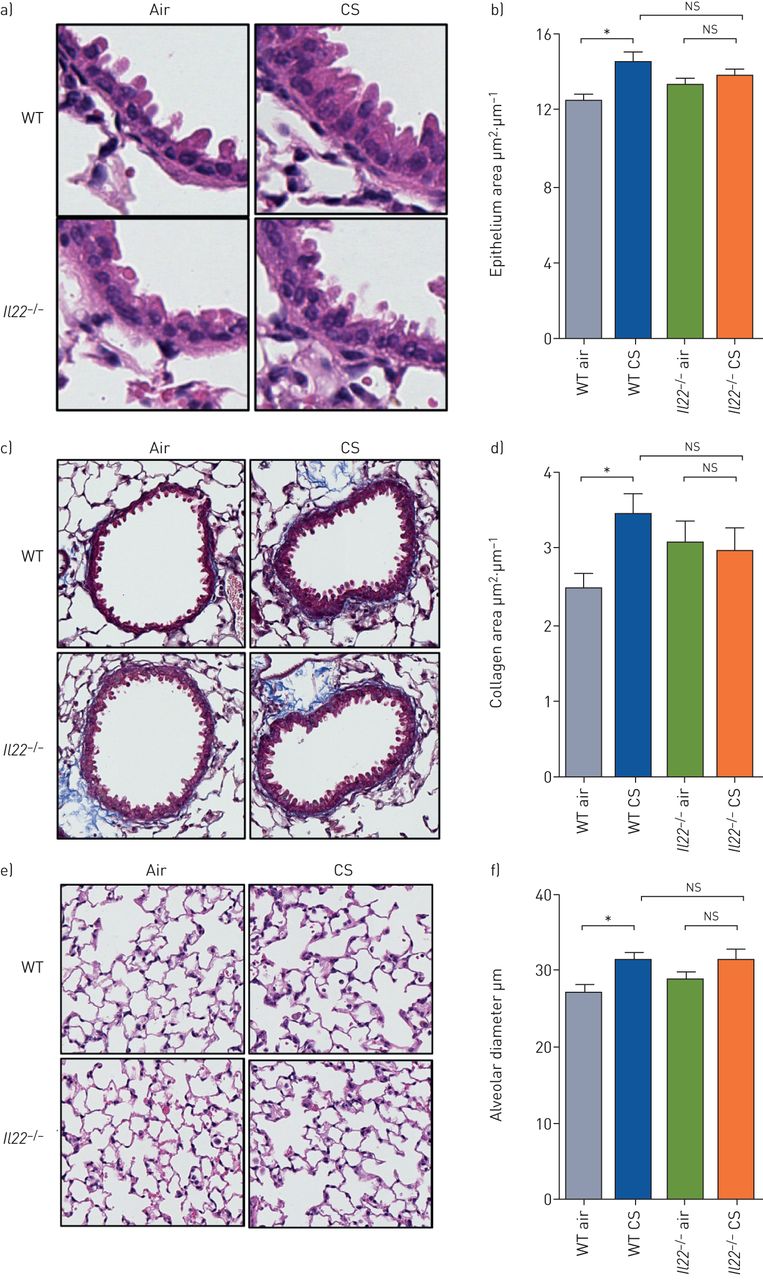
Cigarette smoke (CS)-induced increases in airway epithelial area, collagen deposition and emphysema-like alveolar enlargement do not occur in Il22−/− mice. Wild-type (WT) and interleukin (IL)-22-deficient (Il22−/−) C57BL/6 mice were exposed to normal air or CS for 8 weeks to induce experimental chronic obstructive pulmonary disease. a) Representative images of small airway epithelium; b) small airway epithelial thickness in terms of epithelial cell area (μm2) per basement membrane perimeter (μm); c) representative images of collagen deposition around small airways; d) area of collagen deposition (μm2) per basement membrane perimeter (μm); e) representative images of alveolar structure; f) alveolar diameter (μm). Data are presented as mean±sem, n=6, with another independent experiment showing similar results. The one-way ANOVA with Bonferroni post-test analysed differences between three or more groups. *: p<0.05 compared to normal air-exposed controls; ns: nonsignificant.
We then assessed inflammatory cell numbers in lung tissue sections [12–14, 29]. CS exposure of WT mice significantly increased inflammatory cell numbers in the parenchyma compared to normal air-exposed WT controls (figure 5e–f). CS-exposed Il22−/− mice had increased parenchymal inflammatory cells compared to their normal air-exposed controls. Numbers of parenchymal inflammatory cells were not different between CS-exposed Il22−/− and WT mice.
Next, histopathology was scored according to a set of custom-designed criteria as described previously [30]. CS exposure of WT mice increased histopathology score, which was characterised by increased airway, vascular and parenchymal inflammation (figure 5g–k). CS-exposed Il22−/− mice had increased histopathology, airway, vascular and parenchymal inflammation scores compared to their normal air-exposed controls. Il22−/− mice had a small but significant reduction in total histopathology score, compared to CS-exposed WT controls.
We then profiled the mRNA expression of chemokines and cytokines, other than IL-22, that are involved in neutrophil influx into the lung including chemokine (C-X-C motif) ligand (CXCL)1, CXCL2 and IL-17A [37]. CS-exposure of WT mice resulted in significantly increased Cxcl1, Cxcl2 and Il17a mRNA expression compared to normal air-exposed WT controls with Cxcl1 and Cxcl2 having ∼200-fold greater expression than Il17a (figure 5l–n). CS-exposed Il22−/− mice also had increased expression of Cxcl1 and Il17a, but not Cxcl2, compared to normal air-exposed Il22−/− controls. There was a significant reduction in Cxcl2, but not Cxcl1 or Il17a mRNA expression in CS-exposed Il22−/− mice compared to CS-exposed WT controls. Protein levels of IL-17A, MPO and neutrophil elastase were increased in CS-exposed WT mice, but were unaltered in Il22−/− mice (supplementary figure S11).
CS-induced increases in airway epithelial area, collagen deposition and emphysema-like alveolar enlargement do not occur in Il22−/− mice
We have previously shown that CS-exposed WT mice develop small airway remodelling (increased epithelial area), fibrosis (collagen deposition) and emphysema-like alveolar enlargement after 8 weeks of CS exposure [12–14, 17, 18, 32]. Thus, we determined whether IL-22 contributes to these disease features. In agreement with our previous studies, CS exposure of WT mice increased small airway epithelial cell area compared to normal air-exposed WT controls (figure 6a and b). In contrast, CS-exposed Il22−/− mice had no change in airway epithelial cell area compared to normal air-exposed Il22−/− controls.
CS-exposed WT mice had increased collagen deposition compared to normal air-exposed WT controls (figure 6c and d). However, CS-exposed Il22−/− mice did not have increased collagen deposition compared to Il22−/− normal air-exposed controls.
CS-exposed WT mice had significantly increased alveolar diameter compared to normal air-exposed WT controls (figure 6e and f). CS-exposed Il22−/− mice did not have increased alveolar diameter compared normal air-exposed Il22−/− controls.
As a result of the relatively small differences in airway epithelial area, collagen deposition and alveolar diameter the differences were not statistically different between CS-exposed Il22−/− mice and CS-exposed WT controls.
CS-induced lung function impairment is improved in Il22−/− mice
We assessed the role of IL-22 in CS-induced impairment of lung function, measured in terms of lung volume, airway resistance, TLC, inspiratory capacity, FVC and compliance. CS-exposed WT mice had increases in all of these parameters compared to normal air-exposed WT controls (figure 7a–f). In CS-exposed Il22−/− mice, none of these lung function parameters were significantly different compared to normal air-exposed Il22−/− controls. Again, likely due to small changes in mild-to-moderate experimental COPD, these lung function parameters were not significantly altered in CS-exposed Il22−/− mice compared to CS-exposed WT controls. However, CS-exposed Il22−/− mice had similar lung function to air-exposed WT controls.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cigarette smoke (CS)-induced lung function impairment is improved in Il22−/− mice. Wild-type (WT) and interleukin (IL)-22-deficient (Il22−/−) C57BL/6 mice were exposed to normal air or CS for 8 weeks to induce experimental chronic obstructive pulmonary disease. Lung function was assessed in terms of a) lung volume from pressure–volume loops, b) airway resistance, c) total lung capacity (TLC), d) inspiratory capacity, e) forced vital capacity (FVC) and f) compliance. Data are presented as mean±sem, n=6, with another independent experiment showing similar results. The one-way ANOVA with Bonferroni post-test analysed differences between three or more groups. *: p<0.05 compared to normal air-exposed controls; ns: nonsignificant.
We assessed tissue elastance and found a nonsignificant reduction in CS-exposed WT mice that was not different in Il22−/− mice (supplementary figure S12).
Discussion
Here, we demonstrate that IL-22 plays a previously undefined role in the pathogenesis of CS-induced experimental COPD. IL-22 and its receptors were increased in both human and experimental COPD. We show for the first time, using IL-22 reporter mice, that elevated lung IL-22 levels in experimental COPD result from increased IL-22+ CD4+ T-cells, γδ T-cells, NKT-cells and ILC3s. In addition, we have demonstrated that CS-induced neutrophilic airway inflammation was reduced in Il22−/− mice compared to WT controls. Furthermore, Il22−/− mice did not develop CS-induced airway remodelling and emphysema and had improved lung function that was comparable to normal air-exposed controls. Hence, this study provides new insights into the roles of IL-22 in the pathogenesis of COPD.
The presence or absence of IL-22 may affect resident microbiota. Indeed, we have reviewed the pathogenic roles for gut and lung microbiota in the development of COPD [5, 38, 39]. To minimise the influence of altered microbiota, WT and Il22−/− mice were derived from the same breeding pairs, maintained in the same facility and used experimentally at the same time, and so they would be expected to have very similar microbiomes.
Using pre-existing microarray datasets, we show that IL-22 and IL-22R mRNA expression were increased in airway epithelial cells from patients with mild-to-moderate COPD [20]. However, IL-22 and IL-22R mRNA were unaltered in lung parenchymal cores in severe COPD [22]. Our data are supported by studies that show increased IL-22 protein levels and IL-22+ immune cells in blood, sputum and lung biopsies of COPD patients (reviewed in [8]). However, there are limited reports of IL-22 receptor expression in COPD. Neutrophil proteases have been shown to alter IL-22R-dependent antimicrobial defence in COPD, but there was no change in IL22RA1 mRNA expression in lung tissue or primary cultures of proximal airway epithelial cells from COPD patients compared to healthy controls [9]. IL-10RB and IL-22RA2 have not been assessed in COPD. Consistent with our human data, IL-22 was increased in lung tissue homogenates in experimental COPD after 8 weeks, but not before the development of disease upon 1 week of CS exposure. IL-22 receptor mRNA expression was different between human and mouse. However, at the protein level, IL-22RA1 and RA2 were visually increased in the airway epithelium of CS-exposed mice, which was consistent with changes at the mRNA level in humans. IL-22 receptors were also increased at protein level in human COPD. Collectively, our data show that IL-22 and its receptors are increased in both human and experimental COPD. However, the expression of IL-22 and its receptors is heterogenous and is influenced by tissue location and disease severity.
Given that IL-22 was increased in the lungs in experimental COPD, we used IL-17A and IL-22 dual reporter mice that facilitate the identification of IL-17A- and IL-22-expressing immune cells without ex vivo stimulation or cell fixation. This enables a more accurate determination of the in vivo lung environment. We show for the first time that CS exposure induced IL-22 production from CD4+ T-cells, γδ T-cells, NKT-cells and ILC3s, which are the major cellular sources of IL-22, and all these cell subsets have known roles in COPD pathogenesis [36, 40, 41]. However, the individual contribution of each of these cells to IL-22 production and COPD pathogenesis remains to be fully elucidated, especially in humans.
Previously, the role of IL-22 in the pathogenesis of COPD was largely unknown. We addressed this gap in knowledge using an established mouse model of tightly controlled chronic nose-only CS-induced experimental COPD [12–18]. Our models are representative of a pack-a-day smoker [24]. We have consistently shown that 8 weeks of CS exposure in our models is sufficient to induce the hallmark features of human COPD: chronic inflammation, airway remodelling, emphysema and impaired lung function [12–18]. This 8-week time point was specifically chosen to investigate the underlying pathogenic mechanism(s) during the early stages (GOLD I/II) and identify potential therapeutic targets to halt the progression of COPD.
Using this established model, we show for the first time that IL-22 contributes to COPD pathogenesis independently of infectious exacerbations. Il22−/− mice had reduced airway neutrophils, which was associated with decreased Cxcl2 mRNA expression. CXCL1 and CXCL2 are the mouse orthologues/homologues of human IL-8 and have critical roles in neutrophil influx into the airways following CS-exposure [42]. It has been suggested that improper activation of neutrophils lies at the core of COPD pathology, and mechanisms regulating their function are potential therapeutic targets [43]. However, Il22−/− mice were protected from the increases in MPO or neutrophil elastase levels. Il22−/− mice also had decreased lung tissue inflammation indicated by reduced histopathological score. This is consistent with a previous report showing that administration of recombinant (r)IL-22 into the lung increased tissue inflammation [44].
Additionally, we demonstrate, as we have shown previously, that increases in airway epithelial area, collagen deposition around small airways and emphysema-like alveolar enlargement occur following chronic CS exposure in WT mice [12–18]. Notably, these features did not develop in Il22−/− mice compared to normal air-exposed Il22−/− controls, although the changes were not significant between CS-exposed Il22−/− mice and CS-exposed WT controls. IL-22 is essential for lung epithelial cell repair following influenza virus infection and is implicated in renal fibrosis [45, 46]. Others have shown that mice lacking IL-22 have delayed bacterial clearance and increased alveolar wall thickening and airway remodelling [10]. Administration of rIL-22 with or without acute CS-exposure induced airway epithelial thickening and collagen deposition, although this was not quantified [44].
Our study is the first report on the role of IL-22 in regulating multiple lung function parameters, particularly in models of COPD. We show that Il22−/− mice have improved lung function in terms of lung volumes, airway resistance, TLC, inspiratory capacity, FVC and compliance, comparable to normal air-exposed WT mice. One previous report in an acute CS-exposure model showed increased airway resistance following administration of rIL-22 [44]; however, ours is the first study to assess lung function in Il22−/− mice.
The absence of IL-22 in CS-exposed Il22−/− mice suppressed both airway remodelling and concomitantly the impairment of lung function in experimental COPD. Indeed, CS-exposed Il22−/− mice were protected against increases in epithelial area, collagen deposition and emphysema compared to normal air-exposed controls. Airway remodelling involving epithelial hyperplasia and fibrosis are important in driving resistance to airflow [17, 18]. Emphysema leads to apparent increases in total lung and inspiratory capacity and tissue compliance, which results from the loss of alveolar and parenchymal tissue. In line with the protection against airway remodelling and emphysema-like alveolar enlargement, CS-exposed Il22−/− mice were protected from impaired lung function and changes in airway resistance, total lung and inspiratory capacities and tissue compliance.
In summary, our study demonstrates previously unrecognised roles for IL-22 in COPD pathogenesis. It highlights the potential role of IL-22 in chronic lung diseases, which may be a useful biomarker in the diagnosis and/or prognosis of COPD patients. Furthermore, using a clinically relevant and established model of experimental COPD, our study demonstrates that IL-22 promotes CS-induced pulmonary neutrophilic inflammation, airway remodelling and lung function impairment. However, inhibiting IL-22 may increase the risk of exacerbations due to its central role in pathogen clearance. Therefore, caution in therapeutic approaches targeting IL-22 signalling are required. The relationships between IL-22 and genetic factors, infections/colonisation and phenotypes in COPD remain to be defined.
Supplementary material
Supplementary Material
Please note: supplementary material is not edited by the Editorial Office, and is uploaded as it has been supplied by the author.
Supplementary material ERJ-00174-2018.Supplement
Supplementary figure S1: Gating strategy for lung immune cell subsets. a) CD4+ T cells, b) γδ T cells and NKT cells and c) ILC3. ERJ-00174-2018.Figure_S1
Supplementary figure S2: IL-22 and receptor mRNA in human peripheral lung tissue is unchanged in mild emphysema. Microarray data from peripheral lung tissue of patients with mild emphysema (Accession: GSE8581). a) IL-22, b) IL-22RA1, c) IL-22RA1 and d) IL-10RB. Data are represented as log2 intensity robust multi-array average signals. ERJ-00174-2018.Figure_S2
Supplementary figure S3: No correlation between smoking pack-years and IL-22 or receptor expression. a) IL-22, b) IL-22RA1, c) IL-22RA2. ERJ-00174-2018.Figure_S3
Supplementary figure S4: No change in IL-22 or receptors in bronchial brushings in lung cancer. Microarray data from bronchial brushings in lung cancer (Accession: GSE4115). a) IL-22, b) IL-22RA1, c) IL-10RB. Data are represented as log2 intensity robust multi-array average signals. ERJ-00174-2018.Figure_S4
Supplementary figure S5: No change in IL-22 or receptors in lung tissue in lung cancer. Microarray data from lung tissue in lung cancer (Accession: GSE1650). a) IL-22, b) IL-22RA1, c) IL-10RB. Data are represented as log2 intensity robust multi-array average signals. ERJ-00174-2018.Figure_S5
Supplementary figure S6: Representative images of IL-22 and IL-22 receptor staining in human lung tissue. The four panels are showing the representative images of IL-22 (upper panel), IL-22RA1 (upper middle panel), IL-22RA2 (lower middle panel) and IL-10RB (lower panel) immunohistochemical staining in human peripheral lung tissue. a) and d) Age- and smoke history-matched control smokers with normal lung function. b) and e) Mild-to-moderate stable COPD. Upper lane images show the bronchiolar epithelium whereas lower lanes the alveolar macrophages. Representative images of positive control tissues (tonsils for IL-22, IL-22RA1 and IL-10RB), normal kidney for IL-22RA2 (kindly provided respectively by Prof Stefano Pelucchi and Prof Carmelita Di Gregorio) were stained with primary antibody (c) or with nonspecific immunoglobulin (Ig)G (negative control, f). Total magnification: 1000× (a, b, d, e; bar = 20 μm) or 200× (c, f; bar = 100 μm). ERJ-00174-2018.Figure_S6
Supplementary figure S7: Increased IL-22RA1 protein intensity in the airway epithelium of smokers with COPD. a) IL-22RA1 protein intensity per micrometre (μm) of basement membrane (BM) in non-smokers, healthy smokers without COPD and COPD with or without current smoking separated into GOLD stage 2 and GOLD stage 3–4. b) IL-22RA1 intensity in airway epithelium of non-smokers versus smokers with COPD. c) Representative images of IL-22RA1 positive staining, with red staining in the airway epithelial cells indicating IL-22RA1 positive staining. ERJ-00174-2018.Figure_S7
Supplementary figure S8: IL-22 protein levels are unaltered in the lungs of mice exposed to CS for 1 week. Wild-type (WT) C57BL/6 mice were exposed to normal air or CS for 1 week. IL-22 protein levels in lung homogenates were assessed by ELISA. Data are presented as mean±SEM, n=6, with another independent experiment showing similar results. Two-tailed Mann–Whitney t-test was used to analyse differences between two groups. ERJ-00174-2018.Figure_S8
Supplementary figure S9: Representative images of IL-22RA1 and IL-22RA2 protein in mouse lung tissue sections. Wild-type (WT) C57BL/6 mice were exposed to normal air or CS for 8 weeks. Representative images of negative control (top row), IL-22RA1 and IL-22RA2 staining in mouse lung tissue sections from normal air- (left) and CS-exposed (right) mice. ERJ-00174-2018.Figure_S9
Supplementary figure S10:Representative fluorescence-activated cell sorting (FACS) plots of IL-17A+ and IL-22+ CD4+ T-cells; IL-17A+ and IL-22+ γδ T-cells; IL-17A+ and IL-22+ NKT-cells; and IL-17A+ and IL-22+ ILC3s.ERJ-00174-2018.Figure_S10
Supplementary figure S11: IL-17A, MPO and neutrophil elastase protein levels are increased in experimental COPD, but not in the absence of IL-22. Wild-type (WT) and IL-22-deficient (Il22-/-) C57BL/6 mice were exposed to normal air or CS for 8 weeks to induce experimental COPD. a) IL-17A, b) MPO and c) neutrophil elastase protein levels in lung homogenates. Data are presented as mean±SEM, n=6, with another independent experiment showing similar results. The one-way analysis of variance with Bonferroni post-test analysed differences between 3 or more groups. *: p<0.05 compared to normal air-exposed controls. ns: not significant. ERJ-00174-2018.Figure_S11
Supplementary figure S12: CS induced non-significant reductions in tissue elastance that was not different in (Il22-/- mice. Wild-type (WT) and IL-22-deficient ((Il22-/-) C57BL/6 mice were exposed to normal air or CS for 8 weeks to induce experimental COPD. Lung function was assessed in terms of tissue elastance. Data are presented as mean±SEM, n=6, with another independent experiment showing similar results. The one-way analysis of variance with Bonferroni post-test analysed differences between 3 or more groups. ns: not significant. ERJ-00174-2018.Figure_S12
Acknowledgements
We acknowledge Dale Godfrey (University of Melbourne, Melbourne, Australia) for αGalCer tetramers, Kristy Wheeldon and Nathalie Kiaos for CS exposure of mice, Tegan Moore for assistance with data generation and Jessica Weaver for assistance with Il22−/− and reporter mouse colonies (All at the University of Newcastle, Newcastle, Australia).
Footnotes
This article has supplementary material available from erj.ersjournals.com
Support statement: This study was supported by grants from the National Health and Medical Research Council (NHMRC) of Australia and the Australian Research Council (ARC), The University of Newcastle and Hunter Medical Research Institute. M.R. Starkey was supported by an NHMRC Early Career Research Fellowship and is supported by an ARC Discovery Early Career Researcher Award (DECRA) fellowship. C. Donovan is supported by an NHMRC Early Career Research Fellowship. I.M. Adcock is supported by a Wellcome Trust grant. P.M. Hansbro is supported by an NHMRC Principal Research Fellowship (1 079 187) and by a Brawn Fellowship, Faculty of Health and Medicine, the University of Newcastle. Funding information for this article has been deposited with the Crossref Funder Registry.
Conflict of interest: M.R. Starkey has nothing to disclose.
Conflict of interest: M.W. Plank is a full-time employee of GlaxoSmithKline.
Conflict of interest: P. Casolari has nothing to disclose.
Conflict of interest: A. Papi reports board membership, consultancy, payment for lectures, grants for research and travel expenses reimbursement from Chiesi, AstraZeneca, GlaxoSmithKline, Boehringer Ingelheim, Mundipharma and TEVA, payment for lectures and travel expenses reimbursement from Menarini, Novartis, Zambon and Sanofi, outside the submitted work.
Conflict of interest: S. Pavlidis has nothing to disclose.
Conflict of interest: Y Guo has nothing to disclose.
Conflict of interest: G.J.M. Cameron has nothing to disclose.
Conflict of interest: T.J. Haw has nothing to disclose.
Conflict of interest: A. Tam has nothing to disclose.
Conflict of interest: M. Obeidat has nothing to disclose.
Conflict of interest: C. Donovan has nothing to disclose.
Conflict of interest: N.G. Hansbro has nothing to disclose.
Conflict of interest: D.H. Nguyen has nothing to disclose.
Conflict of interest: P.M. Nair has nothing to disclose.
Conflict of interest: R.Y. Kim has nothing to disclose.
Conflict of interest: J.C. Horvat has nothing to disclose.
Conflict of interest: G.E. Kaiko has nothing to disclose.
Conflict of interest: S.K. Durum has nothing to disclose.
Conflict of interest: P.A. Wark has nothing to disclose.
Conflict of interest: D.D. Sin reports grants from Merck, personal fees for advisory board work from Sanofi-Aventis and Regeneron, grants and personal fees from Boehringer Ingelheim, grants and personal fees for lecturing and advisory board work from AstraZeneca, personal fees for lecturing and advisory board work from Novartis, outside the submitted work.
Conflict of interest: G. Caramori has nothing to disclose.
Conflict of interest: I.M. Adcock has nothing to disclose.
Conflict of interest: P.S. Foster has nothing to disclose.
Conflict of interest: P.M. Hansbro reports funding/consultancies from Pharmaxis, AstraZeneca, Sanofi, Pharmakea, Ausbio, and Allakos outside the submitted work.
- Received January 26, 2018.
- Accepted April 19, 2019.
- The content of this work is not subject to copyright. Design and branding are copyright ©ERS 2019
References










