





Abstract
Growth of fibroblasts from bronchoalveolar lavage fluid (BALF) in patients with systemic sclerosis (SSc) has previously been described. The purpose of the present study was to characterise fibroblasts from BALF and bronchial biopsies from SSc patients with alveolitis and from controls, to analyse fibroblast proliferation, migration, stress fibres and proteoglycan production.
BALF and bronchial biopsies were collected from 10 patients with SSc and alveolitis and from 15 controls.
Outgrowth of fibroblasts was observed from the BALF of four patients, particularly in those with a markedly increased percentage of eosinophils in BALF, but not in any member of the control group. Increased levels of granulocyte-macrophage colony-stimulating factor, correlating with the percentage of eosinophils in BALF, were found in patients when compared with controls. Fibroblasts from BALF showed an elongated, mobile phenotype and increased proteoglycan production compared to the corresponding biopsy fibroblasts.
In conclusion, outgrowth of fibroblasts with an altered phenotype is reported from bronchoalveolar lavage fluid in systemic sclerosis patients with alveolitis and an increased percentage of eosinophils in the bronchoalveolar lavage fluid. These findings indicate a possible role for eosinophil–fibroblast interaction in pulmonary fibrosis in systemic sclerosis.
- Eosinophils
- fibroblasts
- granulocyte-macrophage colony-stimulating factor
- pulmonary fibrosis
- scleroderma
- systemic sclerosis
Systemic sclerosis (scleroderma; SSc) is an autoimmune systemic disease characterised by microangiopathy and fibrosis in skin and internal organs. Pulmonary fibrosis, with or without pulmonary hypertension, is the major cause of mortality 1 and serious morbidity in patients with SSc. Interstitial lung disease, fibrosis and/or alveolitis may be found during post mortem examinations or on high-resolution computed tomography (HRCT) in ∼90% of patients 2. Several studies suggest that alveolitis precedes pulmonary fibrosis in SSc patients and may be diagnosed by HRCT or by bronchoalveolar lavage (BAL), in which increased amounts of inflammatory cells in BAL fluid (BALF) indicate alveolitis 2. Increased percentages of neutrophils and/or eosinophils have been found to be associated with decreased lung diffusing capacity for carbon monoxide (DL,CO) 2. Recently, Bouros et al. 3 reported that higher levels of eosinophils in BALF were associated with increased mortality in SSc patients. High numbers of eosinophils are also found to be associated with poor prognosis in idiopathic pulmonary fibrosis (IPF) 4.
In a previous study by the current authors of SSc patients with short disease duration, the production of versican, biglycan and decorin from skin fibroblasts correlated to changes in skin thickness and echogenicity measured by ultrasound 5. In asthma, a disease characterised by peribronchial fibrosis, increased amounts of several proteoglycans are reported in culture medium from bronchial fibroblasts, where some of these proteoglycans could also be related to hyperreactivity 6. Furthermore, the current authors recently reported that fibroblasts of a thin, elongated phenotype could be cultured from the BALF of asthmatic patients with increased amounts of eosinophils 7. These fibroblasts were activated α-smooth muscle actin (α-SMA) expressing myofibroblasts and showed increased migration and proteoglycan production when compared with corresponding biopsy fibroblasts. Growth of fibroblasts has been noted from the BALF of SSc patients with active lung disease 8. These fibroblasts were reported to have a higher expression of collagen and fibronectin compared to normal, adult lung fibroblasts obtained from autopsy.
Cytokines and growth factors, such as endothelin-1 and transforming growth factor (TGF)-β, released from T-cells and endothelial cells, may have the capacity to activate the fibroblasts to increased proliferation and matrix production 9. Granulocyte-macrophage colony-stimulating factor (GM-CSF) is a growth factor, synthesised and released especially by eosinophils, endothelial cells and fibroblasts. GM-CSF has the capacity to influence proliferation, differentiation and recruitment of eosinophils and is also shown to affect fibroblasts and endothelial cells, potentially resulting in tissue remodelling and angiogenesis 10. Increased levels of GM-CSF in BALF were found in patients with interstitial lung disease with eosinophilia 11.
The purpose of the present study was: 1) to characterise fibroblasts from BALF and from bronchial biopsies obtained from SSc patients with alveolitis and from controls; and 2) to analyse cell content and concentrations of cytokines in the BALF, fibroblast proliferation, migration, stress fibres and proteoglycan production.
PATIENTS AND METHODS
Patients
Included in the present study were 10 patients (four males) aged 29–69 yrs (median 58 yrs) suffering from SSc and alveolitis diagnosed by HRCT (table 1⇓). All patients fulfilled the American College of Rheumatology criteria for SSc: four had diffuse cutaneous SSc 12 and six had limited cutaneous SSc. In the majority of patients the disease duration was short, with median 1.5 yrs and range 0.5–32 yrs. Nine patients out of the 10 were nonsmokers. None of the patients were treated with any putative disease-modifying drug. The control group consisted of 15 healthy, nonsmoking volunteers aged 24–58 yrs. The controls were screened with lung-function tests and a metacholine inhalation provocative dose test causing a 20% fall in forced expiratory volume in one second in order to exclude those with pulmonary disease. Pulmonary function in the patients was assessed by vital capacity (VC), as measured by a dry spirometer, and by DL,CO, as measured by the single-breath method. Both were expressed as per cent predicted value. Skin involvement was assessed by a modified Rodnan skin score.
Clinical characteristics of the systemic sclerosis(SSc) patients
Methods
After informed consent was obtained, BAL was performed with 100–150 mL (5−7×20 mL) of 0.9% sodium chloride solution. The volume of the recovered lavage fluid varied 26.5–70 mL. After the lavage, bronchial biopsies were taken from the central bronchial part of the lung of nine patients and of all controls. To grade the alveolitis, differential cell counts of BALF were performed on cytospin preparations stained by the May-Grünewald-Giemsa method. In total, 400 cells were counted by the same person (M. Wildt). The HRCT scans were scored by a “blinded” observer (S. Ellis) who evaluated the percentage of the lung with ground-glass opacity (GGO) and with reticulation, and also with regions of both GGO and reticulation. In addition, for each of these three patterns the extent of disease was scored in terms of the number of regions involved (min 0; max 6). The scans were split into six regions, three for each lung: below pulmonary venous confluence; between pulmonary venous confluence and carina; and above carina (table 2⇓).
Growth of fibroblasts, recovery, differential counts and high-resolution computed tomography (HRCT) scores in 10 systemic sclerosis (SSc) patients with alveolitis
For the establishment of fibroblast cultures, BALF and biopsies were incubated in 25-cm2 tissue culture flasks in Dulbeco’s modified Eagle’s medium (Sigma Chemicals, St Louis, MO, USA) supplemented with 10% foetal bovine serum (Hyclone, Logan, UT, USA), l-glutamin (2 mM; Sigma Chemicals), amphotericin B (5 μg·mL−1) and gentamicin (50 μg·mL−1). The flasks were stored at 37°C in a humidified cell incubator with 5% CO2 and 95% atmosphere. When confluent, the cells were loosened from the bottom of the flask by treatment with 0.05% trypsin for 70 s and divided into two new bottles.
Concentrations in the BALF of two cytokines were analysed with ELISA; GM-CSF and endothelin (ET)-1. The ELISA methods used were Quantikine (R&D System, Abingdon, UK). The detection limits of the tests were 0.26 pg·mL−1 for GM-CSF and 1.0 pg·mL−1 for ET-1.
Proliferation, migration and morphological characterisation
The growth capacity of fibroblasts was evaluated after 72 h using crystal violet as described previously 7. For the migration assay, 30,000 fibroblasts were seeded and allowed to adhere for 6 h in a cylinder usually used for cloning, which was placed into a chamber. The cylinder was removed and the fibroblasts were allowed to migrate for 48 h. The cells were fixed for 30 min in 1% glutaraldehyde, stained for 2 h in 0.5% crystal violet and washed. The distance for 200 cells was counted by measuring the distance from the border of the removed cylinder. Morphological characterisation was performed with crystal violet 13. Cell shape was recorded as the ratio of the length versus the width of 200 cells.
Stress fibres
For stress fibre analysis, cells were fixed in 4% paraformaldehyde in PBS. After permeabilisation in 0.5% Triton X-100 and blocking in 1% bovine serum albumin (BSA) in PBS, the cells were incubated with Alexa Fluor® 488 phalloidin probe (Molecular Probes, Leiden, the Netherlands) diluted in blocking buffer. A fluorescence microscope was used to examine the cells 13.
Immunostaining of α-SMA
For the immunostaining of α-SMA, monoclonal mouse antibody against human α-SMA (M0851; Dako, Dakopatts AB, Älvsjö, Sweden) was used, followed by Alexa Fluor® 584 goat anti-mouse immunoglobulin G (Molecular Probes) 13.
Western blot
The cell lysate was separated by 4–12% Bis-Tris Gel (Invitrogen, Uppsala, Sweden) in 3-(N-morpholino)propane sulphonic acid running-buffer. The separated proteins were transfered to polyvinylidene fluoride membranes and incubated with primary antibodies against α-SMA (DAKO, Glostrup, Denmark). A secondary horseradish-peroxidase labelled rabbit-antimouse (DAKO) antibody was used and the intensity of the bands were analysed using the Gel-ProTM Analyser software (Media Cybernetics, Silver Spring, MD, USA) 14.
Proteoglycan production
Confluent cultures in passages three to six were labelled 100 μCi·mL−1 medium of (35S)-sulphate for 24 h. The proteoglycans were isolated using ion-exchange gel 6 and further separated into decorin, biglycan, heparan sulphate proteoglycan and versican on sodium dodecyl sulphate polyacrylamide gel electrophoresis 6 and the amounts were calculated per μg of protein using a BSA method.
Statistics
Levels of significant differences between the two groups were calculated using the Mann–Whitney U-test for unpaired observations. The relations between variables were calculated with Spearman′s Rho correlation.
Ethical considerations
The study was reviewed and approved by the local ethical committee (Nr LU 193-01) for patients and controls.
RESULTS
Cell profile of BALF
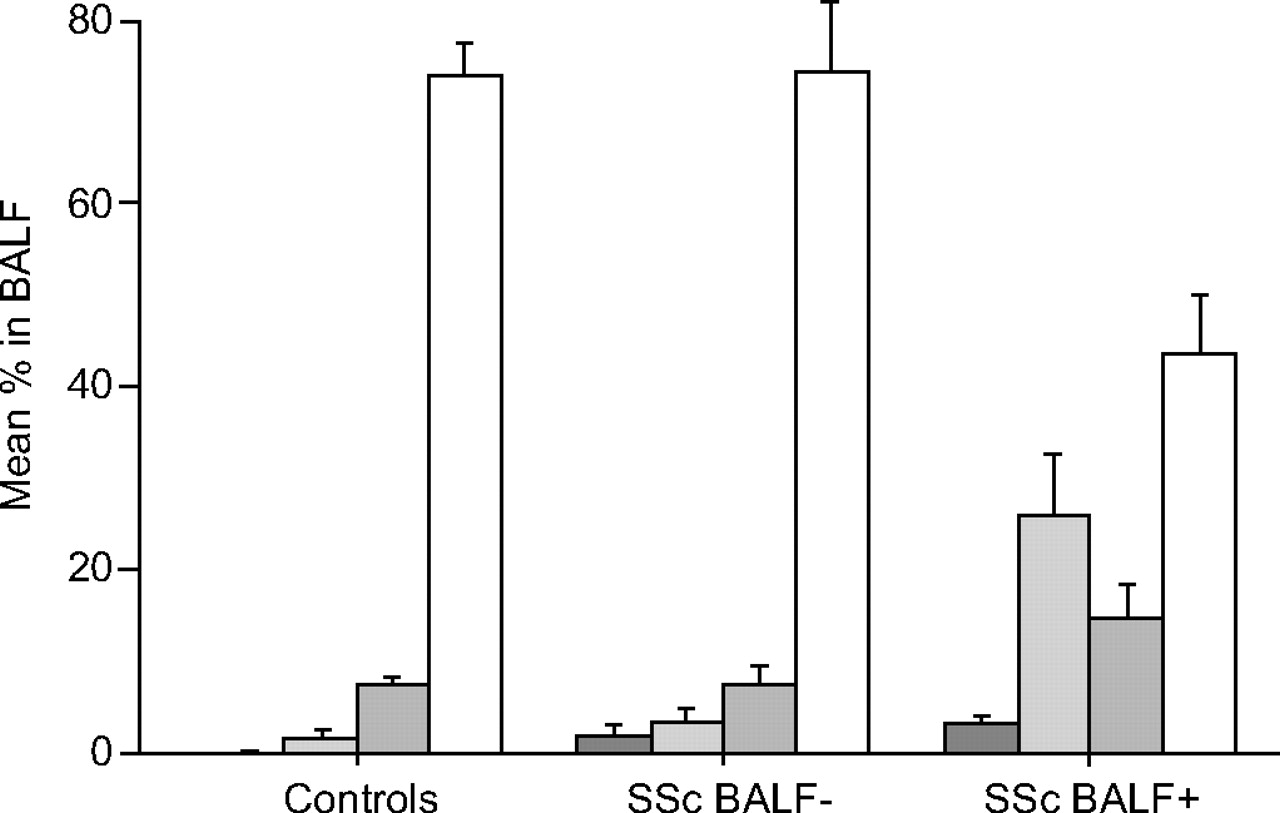
Outgrowth of fibroblasts in SSc patients was found in 50% from biopsies and in 45% from BALF. In contrast, outgrowth of fibroblasts from biopsies from controls was found in 82% but no growth was obtained from BALF from any of the controls (table 2⇑). The total number of cells in BALF in all SSc patients was 0.5–12×10 6 (median 2.5×106; table 2⇑) compared with 0.8–2.8×106 (median 1.05×106) in the controls (p = 0.14). Differential counts of BALF in SSc patients divided according to growth of fibroblasts in BALF and in controls are shown in figure 1⇓. SSc patients had increased percentages of eosinophils (median 2.0%; p<0.01) and neutrophils (median 9.0%; p<0.05) compared with the controls (median 0% and 1%, respectively), whereas no significant difference was found in the percentage of lymphocytes. High percentages of eosinophils and neutrophils were seen, particularly in patients with fibroblast growth in BALF (table 2⇑). There was no correlation between the percentages of neutrophils and eosinophils and VC or DL,CO. The two patients in whom an increased BALF percentage of eosinophils but no fibroblast growth was found were the two most severely affected patients, with markedly decreased pulmonary function despite short disease duration. After assessment, the patients were treated for 1 yr, four with cyclophosphamide, five with azathioprine and one with bosentan. Lung function after 1 yr of therapy is shown in table 1⇑. VC and DL,CO were median 73 and 65% pred before and 85 and 73% pred after therapy in the seven patients evaluable after 1 yr of therapy.

Differential counts (mean and sem) of bronchoalveolar lavage fluid (BALF) in nine systemic sclerosis (SSc) patients with alveolitis and in 13 controls. The patients were divided according to growth (n = 4) or no growth (n = 5) of fibroblasts from BALF. The fluid from one patient was discarded because of contamination. ▒: eosinophils; ░: neutrophils; ▓: lymphocytes; □: macrophages.
Levels of GM-CSF and ET-1 in BALF
Concentrations of GM-CSF in BALF were higher in the patients than in the controls (p<0.02). A correlation was found between the percentage of eosinophils and GM-CSF (fig. 2⇓; p<0.01; rho = 0.81). The percentage of neutrophils was not related to GM-CSF. When GM-CSF was corrected for the percentage of eosinophils, patients with growth of fibroblasts (n = 4) had higher values than patients with no growth (n = 3; p<0.05). Two patients and nine controls had no eosinophils, therefore, the ratio of GM-CSF to eosinophils could not be calculated. Total protein concentration was increased in patients compared to controls, as expected (p<0.01), but was not related to levels of eosinophils. No significant difference was found in BALF concentrations of ET-1; concentrations were found to be median 2.25 (range 0–7.9) pg·mL−1 in patients and 3.0 (1.15–9.5) pg·mL−1 in controls (data not shown).

Correlation between concentration of granulocyte-macrophage colony-stimulating factor (GM-CSF) and percentage of eosinophils in bronchoalveolar lavage fluid (BALF) in nine patients with systemic sclerosis and alveolitis. ○: patients with growth of fibroblasts from BALF; •: patients with no fibroblast growth from BALF.
Characterisation of BALF fibroblasts
SSc fibroblasts cultured from BALF tended to migrate a longer distance (median 477 μm; n = 3) than the biopsy fibroblasts (median 386 μm; fig. 3⇓). The degree of elongation of fibroblasts, as determined by the ratio of cell length to cell width, was calculated. The BALF fibroblasts displayed a thin elongated phenotype (ratio median 6.93; n = 3), when compared to the fibroblasts cultured from the biopsies (ratio median 3.1; n = 3). The fibroblasts were stained to make the stress fibres visible. In BALF fibroblasts, long filaments were seen that were verified to contain long, thick F-actin bundles (fig. 4a⇓), which were not seen in corresponding biopsy fibroblasts (fig. 4b⇓). The presence of the myofibroblast marker α-SMA in intact cell cultures is shown in figure 4c⇓ and d. α-SMA showed no significant difference between BALF- and biopsy-derived fibroblasts when measured by Western blot (median 55 and 57 intensity·μg−1 protein, respectively). A higher 72-h proliferation of BALF fibroblasts (median 2.05; n = 3) was found than of biopsy fibroblasts (1.44; n = 3). The production of biglycan, decorin, versican and perlecan in BALF fibroblasts and corresponding biopsy fibroblasts in an SSc patient is shown in figure 5⇓. For all four proteoglycans, the production was higher in the BALF fibroblasts than in biopsy fibroblasts.
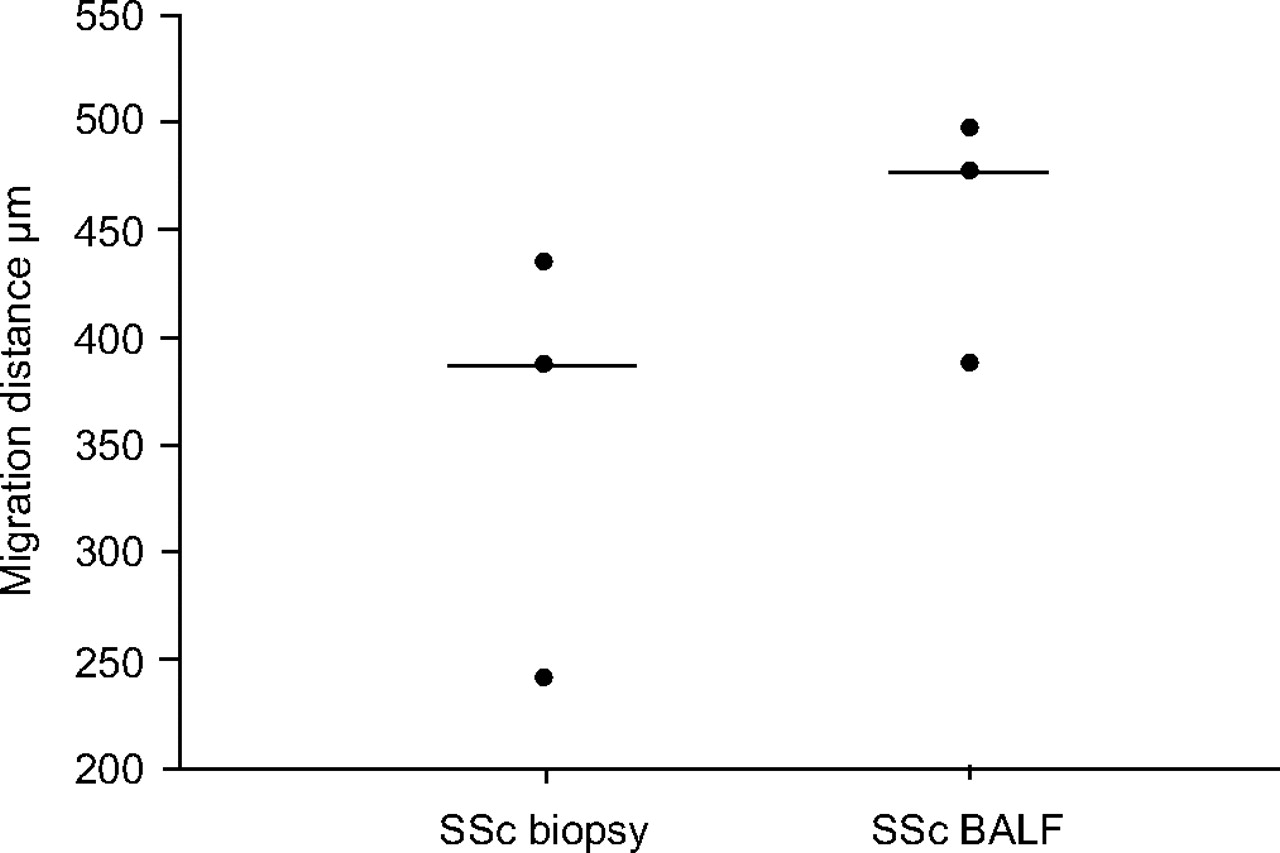
Migration of fibroblasts cultured from bronchoalveolar lavage fluid (BALF) and from bronchial biopsy in systemic sclerosis (SSc) patients with alveolitis. ———: median values.

Stress fibres (bundles of F-actin stained with AlexaFluor® 488 phalloidin) in fibroblasts cultured from a) bronchoalveolar lavage fluid (BALF) and b) bronchial biopsy in a systemic sclerosis patient. α-Smooth muscle actin (α-SMA) in fibroblasts from c) BALF and d) bronchial biopsy with monoclonal mouse antibody against α-SMA followed by AlexaFluor® 584 goat antimouse immunoglobulin G.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Production of proteoglycans, biglycan (•), decorin (○), versican (▵) and perlecan (▾), in fibroblasts cultured from bronchoalveolar lavage (BALF) and from a biopsy from a systemic sclerosis (SSc) patient. DPM: disintegrations·min−1.
Notably, the fibroblasts from the bronchial biopsies of the SSc patients showed no difference from those of the controls with respect to proliferation, migration, cell phenotype or proteoglycan production.
DISCUSSION
The present study aimed to characterise BALF-derived fibroblasts in SSc patients with alveolitis. It shows: 1) that these fibroblasts differ from those obtained by bronchial biopsies in morphology and in their capacity to migrate and produce extracellular matrix proteoglycans; 2) that fibroblast growth from BALF is associated with an increased amount of eosinophils in BALF; and 3) that the amount of eosinophils in BALF correlates positively with levels of GM-CSF in BALF.
To obtain fibroblasts from lung tissue both BAL and bronchial biopsies were performed. Open lung biopsy would have been preferable but is associated with significant morbidity, as is thorascopic biopsy. Since healthy controls were also included in the study this was not an option. Biopsy sampling is a complicated procedure influenced by several factors, such as localisation of the biopsy, which may not be representative of the complete lung. This problem constitutes one of the greatest drawbacks with biopsies. Outgrowth of fibroblasts from biopsies was found in 50% of SSc patients compared with fibroblast outgrowth in 82% of controls. The reason for this discrepancy is not clear but similar results were found in patients with asthma, with outgrowth of fibroblasts in 55% of biopsies (unpublished observations). The current authors speculate that biopsy fibroblasts may represent resident lung fibroblasts and that either the amount of residential fibroblasts or the local milieu in the tissue may have favoured the increased outgrowth of these fibroblasts in healthy controls during cell culture. In similarity with normal lung fibroblasts 15, biopsy fibroblasts in SSc patients stained positively for α-SMA and had features similar to biopsy fibroblasts from the healthy controls.
In the present study, outgrowth of α-SMA-positive fibroblasts from BALF was observed in patients with SSc and alveolitis, but not in controls. These observations confirm previous reports of growth of fibroblasts with human smooth muscle cell differentiation features from BALF in nine out of 19 SSc patients but not in members of a control group 8. BALF fibroblasts were characterised by an elongated, mobile phenotype, with long, extended actin bundles, corresponding to the findings in the asthma patients 7. In support of a more mobile phenotype of BALF fibroblasts compared with biopsy fibroblasts is the recent finding of an increased expression of the small guanosine triphosphatases RhoA and Rac1, which are known to induce stress fibres and focal adhesions both in SSc and asthma patients 14. In addition, an increased production of extracellular matrix proteoglycans in BALF fibroblasts compared with biopsy fibroblasts suggests that BALF fibroblasts may represent one possible source for increased extracellular matrix accumulation and fibrosis, not only in SSc but also in asthma 7. However, BALF fibroblasts from SSc patients differ from those obtained from asthmatic patients in their protein expression pattern of cytoskeletal and scavenger proteins, suggesting additional differences in the pathogenesis and mechanism of fibrosis in SSc 14.
The cellular origin of fibroblasts in BALF is not clear. They may represent differentiated resident fibroblasts from the mesenchyme or result from transdifferentiation of other cell types 16. Finally, they may have developed from haematopoietic stem cells, such as the previously described fibrocytes recruited by different chemotactic substances 17. Fibrocytes are circulating cells that express both haematopoietic markers such as the surface markers CD34, CD45 and CXC chemokine receptor 4 as well as mesenchymal markers such as collagen I and α-SMA. Several factors, e.g. interleukin (IL)-1, TGF-β, chemokines and serum amyloid P (SAP), modulate the appearance and function of fibrocytes 18. In the presence of TGF-β, fibrocytes are differentiated to assume a myofibroblast phenotype expressing α-SMA 19. In a mouse model of allergic asthma, circulating fibrocytes were shown to be recruited into bronchial tissue at allergen exposure and to differentiate into myofibroblasts 20.
In the present study, it was noted that fibroblasts could be cultured from BALF in patients with increased levels of eosinophils, in a similar manner to that previously observed in asthmatic patients 7. Although the link between high eosinophil count in BALF and outgrowth of fibroblasts is not clear, interaction between eosinophils and fibroblasts has been studied in vitro, suggesting a potential pro-fibrotic liaison. First, in co-culture models, eosinophils stimulate fibroblast proliferation, collagen production and lattice contraction, mediated at least in part by TGF-β 21. Secondly, eosinophils store and release TGF-β, which has been shown to induce myofibroblast differentiation in both fibroblasts and epithelial cells 10, 16. Interestingly, GM-CSF derived from human lung fibroblasts enhances survival of eosinophils in vitro 22, and GM-CSF added to fibroblast–eosinophil co-cultures regulates eosinophil density and function 23. Eosinophils are believed to be important for tissue remodelling in asthma and may also play a role in interstitial pulmonary disease 10. With regard to SSc, increased serum levels of eosinophil cationic protein indicating eosinophil activation has been associated with decreased lung function 24. However, the role of eosinophils in the pathogenesis of fibrosis in SSc remains to be elucidated. Thus, it is not known whether eosinophils might favour recruitment and differentiation of fibroblasts and fibrocytes into myofibroblasts or, on the contrary, whether activated fibroblasts release chemokines and cytokines, such as eotaxins and IL-15 which favour recruitment and/or survival of eosinophils.
In the present study, outgrowth of BALF fibroblasts with an altered fibroblast phenotype was observed in patients with both increased eosinophil counts and increased levels of GM-CSF in BALF. Similarly, increased GM-CSF levels were observed in BALF of patients with IPF that was associated with increased eosinophilia 11. These findings may be explained by the effect of GM-CSF on eosinophil survival or eosinophil-specific recruitment 25, which is reflected by the relationship observed between GM-CSF and eosinophils but not neutrophils. Of interest for the pathogenesis of fibrosis in SSc is the notion that BALF neutrophils correlate to the extent of fibrosis on HRCT, while the percentage of eosinophils correlates to the extent of GGO, representing alveolitis which is assumed to precede fibrosis development in SSc 26. The majority of patients in the present study had short disease duration and relatively well-preserved VC (median 70%), indicating that fibrosis development restricting the pulmonary capacity had not taken place yet.
GM-CSF has been studied in the context of fibrosis development especially in asthma and IPF, suggesting a pro-fibrotic effect of GM-CSF 27, e.g. adenoviral vector-mediated gene transfer of GM-CSF to rat lung was reported to induce eosinophilia and fibrosis, and markedly increased TGF-β levels in BALF 28. Until recently GM-CSF has not been studied in the context of SSc. Expression of the GM-CSF receptor-α was confirmed in SSc skin fibroblast in vitro cultures and GM-CSF had an inhibitory effect on collagen I protein and mRNA production, suggesting a rather anti-fibrotic effect 29. In contrast, the previous data showing increased levels of GM-CSF in BALF of patients with SSc associated with increased eosinophilia and an increased outgrowth of proteoglycan-producing fibroblasts with a mobile phenotype suggest a more pro-fibrotic function of GM-CSF. GM-CSF could therefore be involved in the recruitment of inflammatory cells, eosinophils and/or fibrocytes to the pulmonary interstitium. However, further studies are required to address the exact mechanism of GM-CSF on eosinophil and fibroblast interaction and to clarify the role of GM-CSF in the pathogenesis of SSc.
The present study suggests the coexistence of two different types of fibroblasts in the bronchial tree of patients with SSc: “normal” resident lung fibroblasts as obtained by bronchial biopsies and proteoglycan-producing fibroblasts that were associated with high eosinophil counts and increased levels of GM-CSF in BALF . Speculatively, these BALF fibroblasts could be derived from circulating fibrocytes. Surprisingly, no growth was found in the two most severely affected patients with high eosinophil counts, one of whom also had a chronic obstructive pulmonary disease that may have influenced the evaluation of the HRCT scan. Therefore, factors other than GM-CSF may be important for promotion of fibroblasts in BALF in SSc patients, e.g. SAP, which has been reported to be the factor in serum that inhibits fibrocyte differentiation 30. SSc patients had lower serum levels of SAP, suggesting that low levels of SAP may augment the pathological processes leading to fibrosis. The variation within the SSc group was large, possibly reflecting the wide spectrum of disease manifestations in SSc.
In conclusion, systemic sclerosis is a rare disease, which is difficult to diagnose early and has a wide disease spectrum, which hampers studies of the early disease process. Although the material in the present study is small and interpretation of the results is difficult, the results might indicate that eosinophil–fibroblast interaction could have an important role in the tissue remodelling process of pulmonary fibrosis in systemic sclerosis. The increased mortality reported by Bouros et al. 3 in patients with systemic sclerosis and high bronchoalveolar lavage eosinophils and the above findings call for further studies to clarify the role of eosinophils and eosinophil–fibroblast interplay in the pathogenesis of scleroderma pulmonary fibrosis.
Acknowledgments
The authors would like to thank D. Wuttge for valuable discussions on the eosinophil–fibroblast interaction.
- Received November 16, 2005.
- Accepted November 4, 2006.
- © ERS Journals Ltd
References










