





Abstract
Leukotriene (LT)B4 in exhaled breath condensate (EBC) has been reported to be elevated in airway inflammation. The origin of leukotrienes in EBC is, however, not established.
The aims of this study are to measure LTB4 levels in EBC collected in two challenges characterised by a strong neutrophilic airway inflammation and to compare LTB4 levels in EBC with levels in sputum and saliva.
LTB4 and α-amylase were measured in EBC from 34 healthy subjects exposed in a pig confinement building or to a lipopolysaccharide provocation. These markers were also measured in induced sputum in 11 of the subjects. For comparison, LTB4 and α-amylase were measured in saliva from healthy subjects.
Only four out of 102 EBC samples had detectable LTB4 (28–100 pg·mL-1). α-amylase activity was detected in the LTB4-positive samples. In contrast, LTB4 was detected in all examined sputum supernatants in the same study (median 1,190 pg·mL-1). The median LTB4 level in saliva was 469 pg·mL-1.
High levels of leukotriene B4 in saliva and the presence of leukotriene B4 in exhaled breath condensate only when α-amylase was detected, indicate that leukotriene B4 found in exhaled breath condensate is the result of saliva contamination. As leukotriene B4 was consistently present in sputum supernatants, exhaled breath condensate may be inappropriate for monitoring airway leukotriene B4.
Exhaled breath condensate (EBC) has gained considerable attention as a new noninvasive technique for measuring markers of airway inflammation 1. The procedure normally involves tidal breathing for 5−15 min and passage of the expired air through a cold trap where the fluid phase is condensed. A great number of mediators, markers and other molecules have been measured in EBC 1. Following the original description of the method by Sidorenko et al. 2, there are to date >300 papers published on the topic, of which ∼200 are from the past 5 yrs. The method is thus very attractive but, as pointed out in the recent report from a European Respiratory Society Task Force 1, there are methodological issues that remain unresolved. In fact, few experimental studies have satisfactorily explained the processes that determine the transfer of different molecules from the lungs and airways into condensates of exhaled air.
Leukotriene (LT)B4 is a potent chemotactic mediator that may be involved in several inflammatory reactions in the lungs and airways 3, 4. LTB4 is produced predominantly by activated neutrophils, but also by alveolar macrophages and in transcellular reactions involving inflammatory cells and surrounding structural elements 4. In addition to recruiting neutrophils and causing increased microvascular permeability 5, LTB4 may stimulate lymphocyte migration in the lung 6 and constriction of pulmonary blood vessels via indirect mechanisms 7. In comparison with healthy subjects, the LTB4 levels have been reported to be elevated in EBC collected from subjects with asthma 8, cystic fibrosis 9 and chronic obstructive pulmonary disease (COPD) 10.
The primary aim of the present study was to measure the levels of immunoreactive LTB4 in EBC collected in a human model where a pronounced neutrophilic inflammation was induced 11. The model involves a standardised exposure of healthy subjects in a pig house for 3 h. This challenge results in a two- to three-fold increase in bronchial responsiveness to methacholine, and an associated intense airway neutrophilic inflammation. Previous studies have shown a 75-fold increase in the number of neutrophils in bronchoalveolar lavage (BAL) fluid 11 and a two- to 5.5-fold increase in the levels of LTB4 in nasal lavage (NAL) fluid following exposure in a pig house 12, 13. It was therefore considered suitable to use the noninvasive EBC method to measure LTB4 levels in a study in which responses to pig house-dust and lipopolysaccharide (LPS) were to be characterised. For comparison, levels of immunoreactive LTB4 in induced sputum were measured in a subgroup of the subjects. In addition, saliva was collected at baseline from another group of healthy subjects and immunoreactive levels of LTB4 as well as α-amylase activity were measured.
MATERIAL AND METHODS
Subjects
A total of 34 healthy subjects, mean age 38 yrs (range 22–61), participated in the study. None of the subjects had a history of asthma, allergy or other lung diseases and none had suffered from a respiratory tract infection during the week prior to the study.
Saliva was collected from another group of 17 healthy nonsmoking, nonatopic, nonasthmatic volunteers, mean age 31 yrs (22–55).
All subjects gave informed consent to participate in the study, which was approved by the ethical review board at Karolinska Institutet.
Study design
All subjects were exposed to pig house-dust in a pig confinement building while weighing pigs for 3 h and, on a separate day, were provoked with an LPS solution (53 μg) using a dosimeter controlled jet nebuliser (Spira Elektro 2; Intramedic, Bålsta, Sweden). Pig house exposure and LPS challenge were performed in random order separated by 2–5 weeks. Collection of EBC was performed 2–14 days before the first exposure and 6 h after the start of pig house-dust exposure or LPS provocation. Induced sputum was collected at baseline and within 9 h of the start of pig house exposure.
A total of 5 mL of saliva at baseline was obtained from another group of healthy volunteers (n = 17) who were asked to chew on a plastic tube stopper.
Collection of EBC in a siliconised glass condenser
In a preliminary set of experiments, the present authors used the ECoScreen (Jaeger, Wurzburg, Germany) for collection of EBC, but found LTB4 in the EBC only occasionally. It has been reported that the coating of the condenser might be important for the recovery of nonvolatile compounds such as eicosanoids 14. Therefore, in the present study, a silicon-coated glass condenser was used in order to minimise possible binding of the leukotrienes to the coating (fig. 1⇓). The subjects breathed through a standard cardboard mouthpiece (Vitalograph GmbH, Hamburg, Germany) connected to a 32 mm PVC tube with a 90° bend coupled to the inlet of a vertically placed condenser tube. The diameter of the inlet (∼20 mm) was smaller than the PVC tube, allowing a gap which functioned as a saliva trap. The junction was tightened with laboratory film (Parafilm “M”®; Pechiney Plastic Packaging, Chicago, IL, USA). The 20 cm-long condenser was of glass type Allihn, pre-treated with silicon, and refrigerated at 0.5°C by a thermostatic circulator (LKB, Bromma, Sweden). EBC was collected in an ice-chilled 50-mL polypropylene tube. Approximately 5 mL of EBC was collected during 20 min of tidal breathing. Nose-clips were not used. The condensate was immediately aliquoted (0.5 mL·aliquot-1) into 0.75 mL cryotubes (BioStor™ with separate “O”-ring screw caps; National Scientific Supply Co., Inc., Claremont, CA, USA) and stored at -20°C until analysis. The condenser was thoroughly rinsed with distilled water for 3 min after each collection.

Schematic figure of the exhaled breath condenser.
Induced sputum and sputum processing
In a subgroup of 11 healthy nonsmoking subjects, sputum induction and processing were performed as described elsewhere 15, with minor modifications. The subjects were pre-treated with 400 µg inhaled salbutamol (Ventoline®; GlaxoSmithKline AB, Mölndal, Sweden) to inhibit excessive airway constriction. A few subjects could expectorate sputum spontaneously and the rest (the vast majority of the subjects) were induced with an aerosol of isotonic and hypertonic saline, inhaled from an ultrasonic nebuliser (Ultra Neb 2000; DeVilbiss Health Care, Somerset, PA, USA). The nebuliser output was 3 mL·min-1. The concentrations of saline were 0.9, 3, 4 and 5%; the duration of each inhalation concentration was 7 min, followed by forced expiratory volume in one second measurements. After each period of inhalation, the subject was instructed to blow their nose and to rinse their mouth with water to minimise contamination of the sputum sample. The subject was asked to cough deeply and to attempt to expectorate sputum. The sample was considered adequate when it appeared macroscopically to be free from saliva and when it weighed ≥1 g. There was thus no difference in the mean volume of collected sputum before or after pig house-dust exposure.
The colour and weight of the entire sputum were determined. An equal volume of dithiothreitol (DTT; Sputolysin® Reagent; EMD Biosciences, Inc., San Diego, CA, USA) 0.1% was added to the sample for a final concentration of 0.05% DTT and rocked for 15–25 min in a 37°C water bath for homogenisation and to dissociate disulphide bonds. The sample was centrifuged (10 min at 280×g) and the supernatant dispensed into several aliquots, which were kept at -70°C until analysis.
Saliva sampling and processing
Subjects were asked to wash their mouth with water before saliva collection. The sample was centrifuged at 1,500×g for 15 min and the supernatant was stored at -70°C until analysis. LTB4 levels and α-amylase activity were measured.
Measurements of immunoreactive LTB4
LTB4 measurements were performed with an LTB4-specific enzyme immunoassay (EIA), as described by the manufacturer (Cayman Chemical Company, Ann Arbor, MI, USA), in duplicates of serially diluted samples. The assay detection limit was 3.9 pg·mL-1. In all samples, the intra-assay variation was <10% and interassay variation <12%. Analysis was performed within 3−5 months of EBC collection, and within 4 months of saliva collection.
Immunoreactive LTB4 levels were measured in the sputum supernatant as described by the manufacturer (Cayman Chemical Company), with the modification that DTT of the same concentration as in the sputum supernatant (0.05% DTT) was added to the standard curve and EIA buffer. In the modified assay, the intra-assay variation was 1.9% and interassay variation 4.6%.
Measurements of α-amylase activity
Analysis of α-amylase activity in undiluted EBC samples was performed within 3−5 months after EBC collection. The analysis was performed after modification of the method of Calzyme Laboratories, Inc. 16. Thus, 50 μL of undiluted EBC samples in a double set, 50 μL saliva samples in serial dilution 1:100−1:12,800 in a double set, 50 μL sputum supernatant samples in serial dilution 1−1:400, 50 μL α-amylase standard (Sigma-Aldrich Inc., St. Louis, MO, USA) in serial dilution 10−0.078 U·mL-1 (the limit of detection was thus 0.078 U·mL-1), or 50 µL 20 mM glucose solution in serial dilution 20−0.15 mM, respectively, were added on a 96-well plate (Nunc A/S, Roskilde, Denmark). Starch solution (1%) in a volume of 50 µL was added to all wells, except those containing glucose and one set of samples (the difference between samples with and without added starch accounted for the endogenous blank). The plate was left at room temperature (RT) for 10 min, followed by the addition of 100 µL of colour reagent (a mixture of dinitrosalicylic acid and sodium-potassium-tartrate tetra-hydrate, both Sigma-Aldrich Inc.). The plate was sealed (Plate sealer; In vitro AB, Stockholm, Sweden) and placed in a boiling water bath for 12 min. Finally, the plate was read at RT in a spectrophotometer (EL808 Ultra Microplate Reader; Bio-Tek Instruments Inc., Winooski, VT, USA) at 540 nm and the amount of α-amylase was calculated. The intra-assay variability was 2.8% and interassay variability was 14.8% (n = 5).
Stability test
The stability of LTB4 in EBC and in saliva was studied. Aliquots of EBC were supplied with 50 pg·mL-1 of synthetic LTB4 (Cayman Chemical Company) and with saliva (1%). The samples were stored at RT. Aliquots were kept at -70°C immediately after LTB4 addition, as well as after 1, 2, 7 and 24 h. Aliquots were also stored at -20°C for a long-term stability study, and LTB4 immunoreactivity was measured at regular intervals for 100 days.
Spiking EBC with increasing amounts of saliva
EBC (5 mL) and saliva (5 mL) were collected from one healthy subject. Increasing amounts of saliva were added to EBC aliquots. Immunoreactive LTB4, as well as α-amylase activity, were measured as previously described.
Statistical analysis
Data are expressed as median (range), unless otherwise stated. Statistical calculations were performed using the Spearman rank test.
RESULTS
Immunoreactive LTB4 in EBC of healthy subjects exposed in a pig house and to LPS
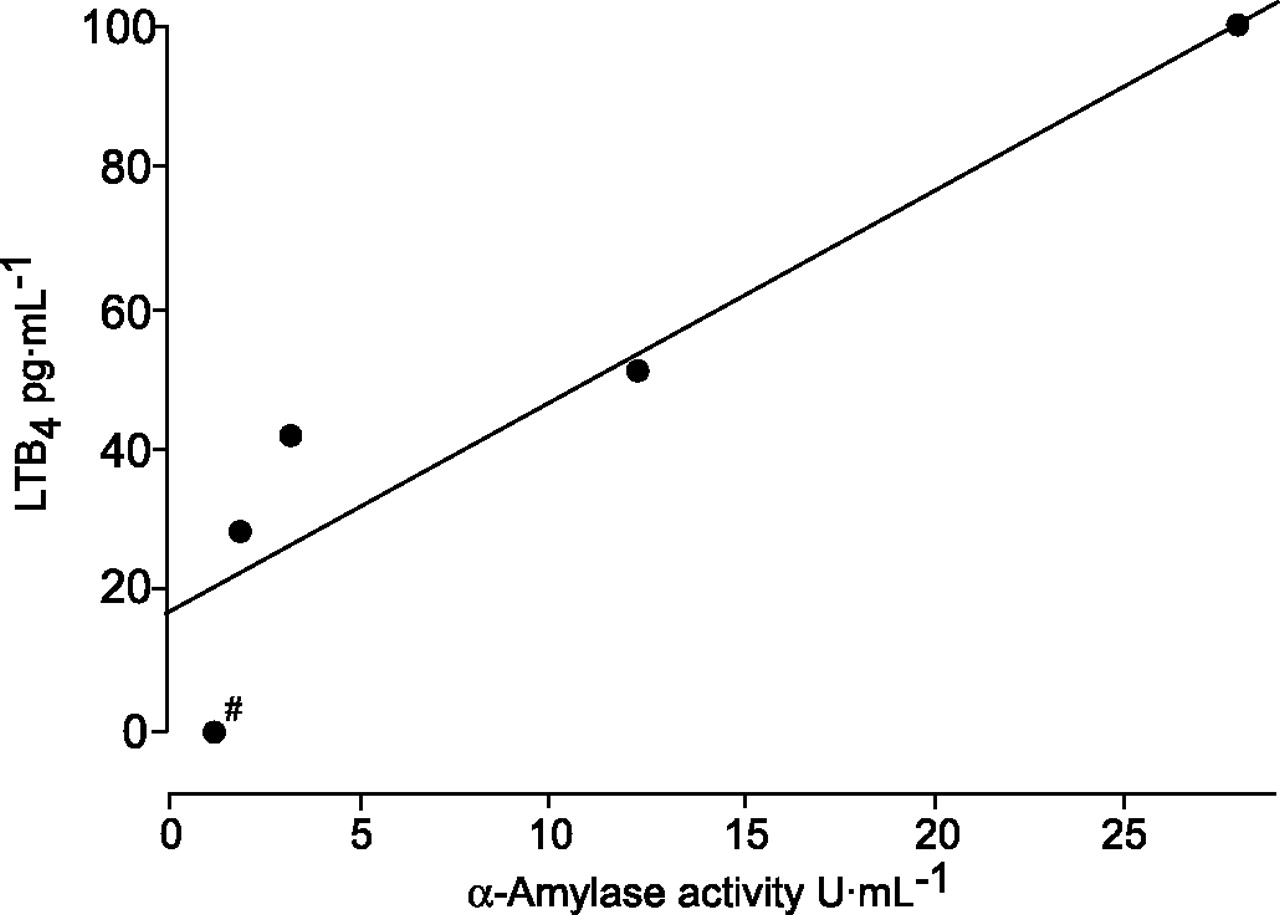
EBCs (n = 102) from 34 subjects examined on three occasions, i.e. before and after exposure in a swine house and after LPS provocation (in a crossover study), were examined for LTB4 immunoreactivity. Irrespective of when they were collected, only four out of the 102 EBC samples tested showed immunoreactive LTB4 levels above the detection limit (3.9 pg·mL-1), with levels ranging 28−100 pg·mL-1 (median 46.5). α-amylase activity was detected in only five of 102 samples: the four samples with detectable LTB4 and in an additional sample (low α-amylase activity) with LTB4 below the detection limit (fig. 2⇓). In these five samples, the α-amylase activity ranged 1.2−28.0 U·mL-1 (3.2 U·mL-1). There was a strong correlation between immunoreactive LTB4 levels and α-amylase activity in the EBC samples (rs = 1.00; p = 0.017; n = 5; fig. 2⇓).

Five exhaled breath condensate (EBC) samples (out of 102) in which leukotriene (LT)B4 or α-amylase was found to be above the detection limit. Levels of immunoreactive LTB4 and α-amylase activity in EBC were significantly correlated (rs = 1.0; n = 5). #: below detection limit for LTB4.
Immunoreactive LTB4 and α-amylase activity in sputum of healthy subjects exposed in a pig house
In the 11 healthy nonsmokers who were exposed in a pig house for 3 h, the median level of immunoreactive LTB4 in the sputum supernatant (pre- and post-exposure) was 1,190 pg·mL-1 (range 193–5,290; n = 22; fig. 3⇓). The median α-amylase activity was 22.3 U·mL-1 (pre- and post-exposure; range 0.9–257.7 U·mL-1; n = 22). Only one EBC sample from these subjects contained detectable LTB4 levels (fig. 3⇓).

Levels of immunoreactive leukotriene (LT)B4 in exhaled breath condensate (EBC) and induced sputum in 11 healthy subjects before and after exposure in a pig house (n = 22). The median level of immunoreactive LTB4 in the sputum supernatants was 1,190 pg·mL-1 (range 193–5,290). Immunoreactive LTB4 could only be detected in one EBC sample collected from the same subjects up to 2 h before sputum induction. The EBC sample had a value of 28 pg·mL-1, whereas the corresponding value in the sputum supernatant was 281 pg·mL-1. ----: detection limit. #: samples with LTB4 levels below the detection limit.
Stability of LTB4 in EBC and saliva
As measurements of immunoreactive LTB4 were performed after storage in the freezer at -20°C, the stability of immunoreactive LTB4 under conditions relevant to the study was established. Immunoreactive LTB4 levels were stable for 24 h at RT in EBC and saliva (fig. 4a⇓), and for >3 months when kept at -20°C (fig. 4b⇓).

Stability of immunoreactive leukotriene (LT)B4. a) Immunoreactive LTB4 levels were stable for 24 h at room temperature in exhaled breath condensate (EBC) and saliva. b) Immunoreactive LTB4 levels were stable for >3 months when kept at -20°C. ▾: EBC supplied with 50 pg·mL-1 LTB4; ▿: EBC supplied with 50 pg·mL-1 LTB4 and saliva (1/100 volume/volume (v/v)); □: saliva (1/10 v/v); ○: EBC; ▪: saliva (1/100 v/v).
Immunoreactive LTB4 levels and α-amylase activity in saliva
In order to confirm that saliva contamination may add LTB4 to EBC samples, saliva was collected from 17 healthy volunteers. Immunoreactive LTB4 levels in saliva were 469 pg·mL-1 (140–1,358). α-Amylase activity in the same samples was 426 U·mL-1 (147–807).
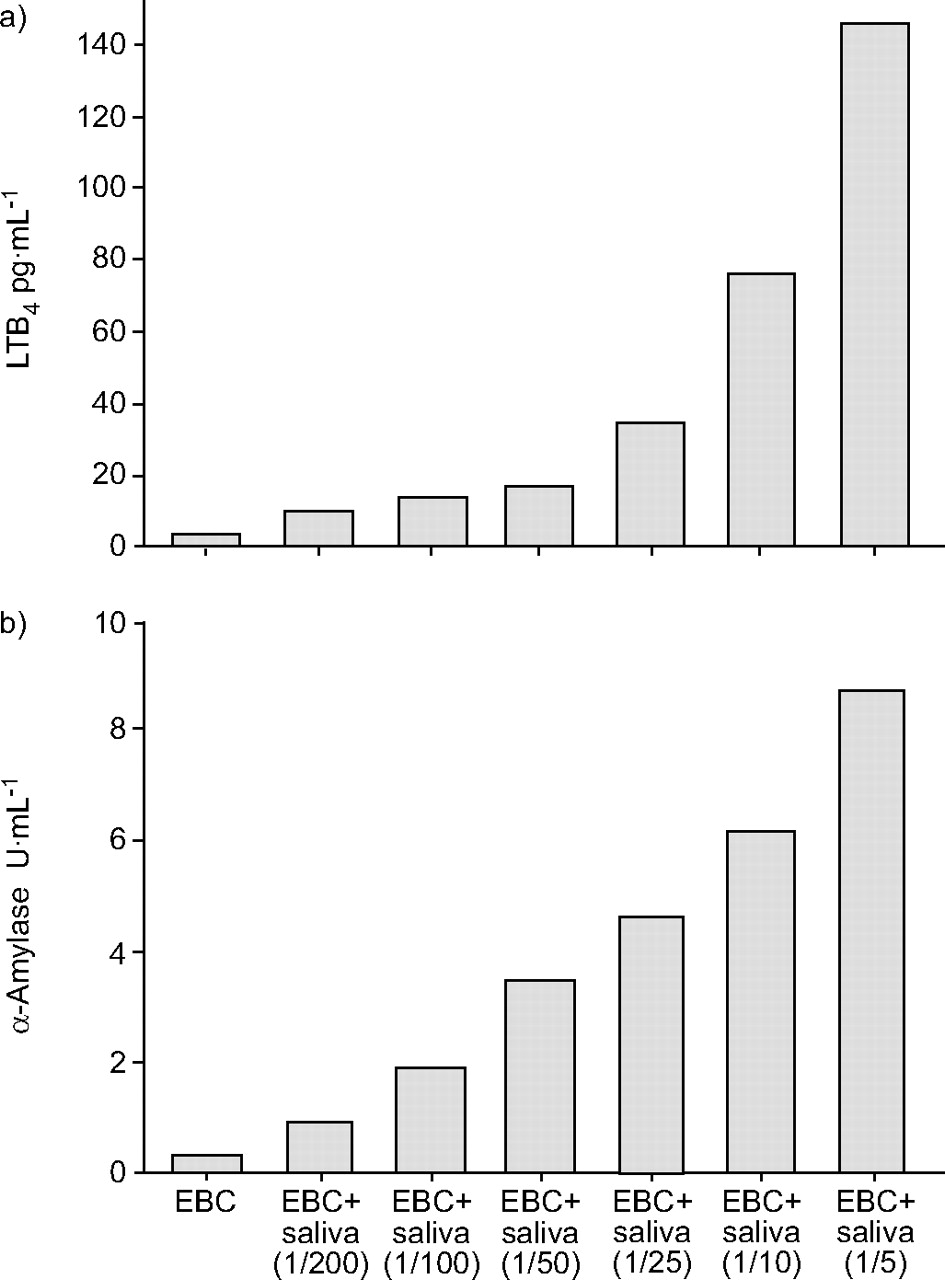
Finally, spiking EBC with increasing amounts of saliva produced consistent parallel increments in LTB4 levels and α-amylase activity, respectively (fig. 5⇓).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
The figure shows increasing amounts of saliva (volume/volume) added to exhaled breath condensate (EBC). a) Immunoreactive leukotriene (LT)B4 and b) α-amylase activity.
DISCUSSION
The authors of the present study were unable to consistently detect LTB4 in 102 samples of EBC collected on three occasions from 34 volunteers. High levels of LTB4 were measured in sputum supernatants collected from 11 subjects before and after pig house-dust exposure in the same study. In a parallel experiment, LTB4 levels in saliva were found to be high (140–1,358 pg·mL-1) in healthy volunteers. Through use of a sensitive method for measurement of α-amylase in EBC, it was found that all samples that were positive for LTB4 also contained α-amylase. Therefore, it was concluded that saliva is a possible source of LTB4 in EBC.
LTB4 levels in EBC have been reported to be increased in several respiratory diseases such as asthma, COPD 17 and cystic fibrosis 9. The levels reported are generally in the 10–130 pg·mL-1 concentration range, although, in more recent publications, the data are expressed as amounts of LTB4 obtained during a defined time period of tidal breathing 18. The vast majority of published papers report on LTB4-like immunoreactivity (LTB4-IR) by the use of EIA measurements, but a recent paper by Montuschi et al. 18 validated the measurements of LTB4 with mass spectrometry. There is, thus, no doubt that LTB4 may be present in EBC.
However, the question is: what is the source of LTB4 in EBC? It is conceivable that material from the pharynx, the nasal cavity and the mouth may contaminate air from the lower airways before exhalation. Many investigators who measure leukotrienes in EBC have considered this and therefore tested EBC for the presence of α-amylase, an enzyme produced by salivary glands 19. An α-amylase activity below the detection limit of the particular assay used, ∼20 U·mL-1 in EBC 20, has been taken as evidence that there is no significant salivary contamination of the EBC and that the EBC sample exclusively contains material from the lower airways. The results of the present study, however, call into question the capacity of measurement of α-amylase in EBC with a standard assay to exclude saliva contamination of the EBC sample.
In the present study, levels of LTB4 (140–1,358 pg·mL-1) and α-amylase activity (147–807 U·mL-1) in saliva were measured in a group of healthy volunteers (n = 17). Also using another method, α-amylase activity in saliva has been found to be highly variable (3–423 U·mL-1; n = 75) 21. Thus, the amylase method originally developed for measurement in saliva is not adequate to detect saliva contamination in EBC. Therefore, a more sensitive assay was developed with a detection limit of 0.078 U·mL-1. As shown by the spiking experiments in the present study (fig. 5⇑), saliva addition ≥0.5–1% gives rise to increased α-amylase and LTB4 in EBC samples. This effect rises strikingly with increased saliva addition.
In the present study, EBC was collected from a group of healthy volunteers exposed in a pig house. This exposure induces an intense neutrophilic inflammation, with 70- and 20-fold increment of neutrophils in BAL and NAL fluid, respectively 22. The levels of LTB4 in NAL have been found to increase 2.5−≥5-fold after exposure 12, 13 and the level of interleukin-8 ∼7−8-fold following exposure in a pig barn 12, 22. The 11 healthy nonsmoking volunteers in the present study had high levels of LTB4 in sputum samples (∼1,000 pg·mL-1). Nevertheless, LTB4-IR was only detected in one of the EBC samples collected from these subjects.
A stability test revealed that the levels of immunoreactive LTB4 in EBC are stable for 24 h at RT and ≥3 months at -20°C. The current authors also investigated whether saliva in EBC caused a difference in LTB4 stability. No difference was found between samples supplied with only LTB4 and samples supplied with LTB4 and saliva (1%). Thus, the storage of EBC samples for some months until analysis does not appear to alter the outcome of the LTB4 assay. Kostikas et al. 17 claimed that the immunoreactivity of leukotrienes in EBC decreases after 3 weeks of storage. However, in that particular study, LTB4 levels dropped to 92% of the original immunoreactivity after the third week, which was within the range of the reported interassay variability of 15%.
There is data supporting the hypothesis that turbulent flow enhances recovery of some compounds in EBC 1. The differences in LTB4 levels in EBC between different patient groups might also reflect events in the upper airways and oral cavity. LTB4 is the major leukotriene produced in the nasal mucosa 23, and there is a significant comorbidity between asthma and rhinitis. Cap et al. 24 reported higher leukotriene levels in EBC in patients with seasonal allergic rhinitis as compared with controls. Another factor could be that most patients with COPD are smokers and smokers have increased prevalence of periodontitis. Interestingly, in the aforementioned study by Kostikas et al. 17, the levels of LTB4 in the EBC of smokers without COPD were found to be high. There are also indications from measurements of other substances that the oropharyngeal tract contributes to the contents of EBC. In fact, a recent study by Marteus et al. 25 showed that nitrite from saliva is an important contaminant of EBC collections.
The present data support the hypothesis that LTB4 in EBC from healthy volunteers derives predominantly from saliva. Low or undetectable levels of LTB4 in EBC from healthy controls have been reported with elevated levels in subjects with different lung diseases. LTB4 was not detected in ∼100 EBC samples in the present study, whereas high levels of LTB4 were measured in induced sputum samples collected before and after the exposure. As in preliminary studies (see Methods) the present authors had also used the ECoScreen condenser, with similar negative results, the present data are not likely to be related to the collection system. These findings, therefore, call into question the appropriateness of EBC as a relevant sample to assess mediators in the lower airways, at least in the case of LTB4. Conversely, the theory that LTB4 could be formed in the lower part of the airways is not questioned, since sputum samples showed a high neutrophil content 26 and LTB4 concentration after exposure in a pig house. However, as Marteus et al. 25 have concluded, the aerosol particles that are supposed to transport LTB4 from the airways may to a much greater extent remain in the exhaled air when formed in the very proximal (including the oropharyngeal tract) rather than the peripheral airways.
In conclusion, the findings of the present study support the hypothesis that the presence of leukotriene B4 in exhaled breath condensate is most likely to be due to salivary contamination. The differences in levels of leukotriene B4 in exhaled breath condensate that have been reported between different groups of patients, or between patients and healthy controls, are generally relatively modest and may, as discussed, be explained by other mechanisms than true differences in recovery of leukotriene B4 from the lower airways. It remains to be seen whether the present conclusion applies only to measurement of leukotriene B4 in exhaled breath condensate, but it is hypothesised that the salivary origin of material in exhaled breath condensate is likely to be a major confounding factor for all compounds that are secreted in relatively high quantities in the saliva. In order to test this possibility, future measurements in exhaled breath condensate need to include a sensitive α-amylase assay and measurements of the particular compound studied in saliva. It is apparent that many publications reporting levels of leukotrienes in exhaled breath condensate do not present values on α-amylase activity.
Acknowledgments
The authors wish to thank A. Ek, M. Olsson, K. Sahlander, I. von Scheele and K. Strandberg for valuable contributions. The authors also wish to thank P. Gerde for helpful discussions. H. Marteus and K. Alving are also gratefully acknowledged for helpful discussions, for kindly lending the ECoScreen equipment and for introducing the authors to the method.
- Received December 26, 2005.
- Accepted August 17, 2006.
- © ERS Journals Ltd
References










