





Abstract
The aim of the study was to examine the effects of a mucolytic drug, carbocisteine, on rhinovirus (RV) infection in the airways.
Human tracheal epithelial cells were infected with a major-group RV, RV14.
RV14 infection increased virus titres and the cytokine content of supernatants. Carbocisteine reduced supernatant virus titres, the amount of RV14 RNA in cells, cell susceptibility to RV infection and supernatant cytokine concentrations, including interleukin (IL)-6 and IL-8, after RV14 infection. Carbocisteine reduced the expression of mRNA encoding intercellular adhesion molecule (ICAM)-1, the receptor for the major group of RVs. It also reduced the supernatant concentration of a soluble form of ICAM-1, the number and fluorescence intensity of acidic endosomes in the cells before RV infection, and nuclear factor-κB activation by RV14. Carbocisteine also reduced the supernatant virus titres of the minor group RV, RV2, although carbocisteine did not reduce the expression of mRNA encoding a low density lipoprotein receptor, the receptor for RV2.
These results suggest that carbocisteine inhibits rhinovirus 2 infection by blocking rhinovirus RNA entry into the endosomes, and inhibits rhinovirus 14 infection by the same mechanism as well as by reducing intercellular adhesion molecule-1 levels. Carbocisteine may modulate airway inflammation by reducing the production of cytokines in rhinovirus infection.
Rhinoviruses (RVs) are the major cause of the common cold and the most common acute infectious illnesses in humans 1. RVs are also associated with acute exacerbations of bronchial asthma 2 and chronic obstructive pulmonary disease (COPD) 3. Several mechanisms of action have been proposed, and the manifestations of RV-induced pathogenesis are thought to be the result of virus-induced mediators of inflammation 3–5.
RV infection induces the production of cytokines, including interleukin (IL)-1, IL-6 and IL-8 5–7. These cytokines exert pro-inflammatory effects 8 and may be related to the pathogenesis of RV infections. Mucolytic or mucoactive drugs, such as l-carbocisteine or carbocysteine lysine salt monohydrate (SCMC-Lys), are used clinically in patients with COPD and bronchial asthma in various countries, including Japan and Italy 9. SCMC-Lys reduces the concentration of IL-6 in the breath condensate of acute COPD patients 10, suggesting anti-inflammatory effects of SCMC-Lys in COPD. SCMC-Lys 11 and S-carboxymethylcysteine (SCMC) 12 also reduce the number of inflammatory cells in airways after exposure to cigarette smoke or sulphur dioxide in rats, and inhibit neutrophil activation 13. These findings are suggestive of anti-inflammatory effects of mucolytic drugs, including carbocysteine. However, the effects of carbocisteine on cytokine production in airway epithelial cells with RV infections have not been studied.
Recent reports have revealed that the major group of RVs enter the cytoplasm of infected cells after binding to their receptor intercellular adhesion molecule (ICAM)-1 14, 15. The entry of the RNA of a major group RV, RV14, into the cytoplasm of infected cells is thought to be mediated by destabilisation from receptor binding and endosomal acidification 15. Macrolide antibiotics, such as bafilomycin 16, 17 and erythromycin 7, inhibit infection by the major group of RVs via a reduction in ICAM-1 expression 7, 17 and an increase in endosomal pH 7, 16. Glucocorticoid also inhibits RV14 infection via reduction of ICAM-1 expression 18. Airway inflammation induced by viral infections, including RV infections, is associated with exacerbations of COPD 3, 4. SCMC-Lys prevents acute exacerbations of COPD 9. However, the mechanisms, other than anti-inflammatory effects in COPD 10, are still uncertain. Conversely, N-acetylcysteine, another mucolytic drug, reduces the expression of ICAM-1 in the lung 19. Therefore, it is conceivable that carbocisteine may modulate the function of airway epithelial cells, including expression of ICAM-1, and may inhibit RV infection. However, the effects of carbocisteine on RV infection have not been studied.
Therefore, the effects of carbocisteine on RV infection in human airway epithelial cells were studied. The effects of carbocisteine on the production of ICAM-1 and cytokines, and on endosomal pH, were also examined in order to clarify the mechanisms responsible for the inhibition of RV infection.
MATERIALS AND METHODS
Patient characteristics
Tracheae for cell culture were obtained after death from 25 patients (mean age 64±2 yrs; 10 female, 15 male) under a protocol approved by the Ethics Committee of Tokohu University School of Medicine. The causes of death included acute myocardial infarction (n = 8), malignant tumour other than lung cancer (n = 8), cerebral bleeding (n = 3), rupture of an aortic aneurysm (n = 2), renal failure (n = 2), congestive heart failure (n = 1) and malignant lymphoma (n = 1).
Human tracheal epithelial cell culture
Isolation and culture of the human tracheal surface epithelial cells were performed as described previously 7, 17.
Viral stocks and detection and titration of viruses
Stocks of a minor group RV, RV2, and a major group RV, RV14, were prepared from patients with common colds by infecting human embryonic fibroblast cells as described previously 7, 17. Detection and titration of RVs were performed by observing the cytopathic effects of the viruses on the fibroblast cells using previously described methods 7, 17, and the amount of specimen required to infect 50% of the fibroblast cells (50% tissue culture infective dose (TCID50)) was determined.
Detection and quantification of rhinovirus RNA
Detection and quantification of RV14 RNA in human tracheal epithelial cells were performed by RT-PCR as previously described 7, 17. In addition, in order to quantify RV14 RNA and reduced glyceraldehye-3-phosphate dehydrogenase mRNA expression in the cells after RV infection, real-time quantitative RT-PCR, using the Taqman technique (Roche Molecular Diagnostic Systems, Foster, CA, USA), was performed as previously described 17, 20–22. The program PrimerExpress (Applied Biosystems, Foster, CA, USA) was used to design the probe and primers based on guidelines for the optimal performance of the PCR 7, 17.
Measurement of lactate dehydrogenase concentration
The amount of lactate dehydrogenase (LDH) in the culture supernatants was measured using the method described by Amador et al. 23.
Effects of carbocisteine on viral infection
In order to examine the effects of carbocisteine on viral titres and the cytokine content of supernatants, and the mRNA expression of ICAM-1, the receptor for the major group of RVs 14, and the low-density lipoprotein (LDL) receptor, the receptor for the minor group of RVs 24, and RV14 RNA expression, the cells were treated with 10 μM carbocisteine or vehicle (PBS) beginning 3 days before RV infection and continuing until the end of the experiments 17. This time-frame was chosen because the maximum concentrations of SCMC in the serum become >10 μM after oral ingestion of 1,500 mg SCMC 25. The cells were then exposed to RV2 (1×105 TCID50·mL-1), RV14 (1×105 TCID50·mL-1) or vehicle (Eagle’s minimum essential medium) for 60 min and cultured at 33°C with rolling, as described previously 7, 17.
Study protocol
In order to measure the time course of viral release during the first 24 h, four separate cultures from the same trachea were used, and the results calculated from seven different tracheae. The culture supernatants were collected at either 1, 6, 12 or 24 h after RV14 infection. In order to measure viral titre during the 24–48 h after RV infection, supernatants were also collected at 48 h after RV infection. The viral content of the supernatant was expressed in TCID50 units per millilitre.
In order to examine the concentration-dependent effects of carbocisteine on RV infection, cells were treated with carbocisteine at concentrations ranging 0.01–30 μM.
The effects of carbocisteine on susceptibility to RV14 infection were evaluated as previously described 6, 7, using epithelial cells pre-treated with carbocisteine (10 μM, 3 days) or vehicle (PBS, 3 days). The cells were then exposed to serial 10-fold dilutions of RV14 or vehicle (Eagle’s minimum essential medium) for 1 h at 33°C. The presence of RV14 in the supernatants collected 1–3 days after infection was determined using the human embryonic fibroblast cell assay, described above, in order to assess whether infection occurred at each RV dose used.
Measurement of intercellular adhesion molecule-1 and low-density lipoprotein receptor expression
The mRNAs of ICAM-1 and LDL receptor were examined using real-time RT-PCR analysis as previously described 7. In addition, concentrations of a soluble form of ICAM-1 (sICAM-1) were measured in culture supernatants using an enzyme immunoassay.
Effects of carbocisteine on cytokine production
IL-1β, IL-6, IL-8 and tumour necrosis factor (TNF)-α were measured in culture supernatants using specific ELISAs 7.
Measurement of changes in acidic endosome distribution
The fluorescence intensity of acidic endosomes in the cells was measured, as previously described, using a dye, LysoSensor DND-189 (Molecular Probes, Eugene, OR, USA) 7, 17, from 100 s before to 300 s after the treatment with carbocisteine (10 μM) or vehicle (PBS).
Isolation of nuclear extracts and electrophoretic mobility shift assays
The extraction of nuclei and electrophoretic mobility shift assays were performed as previously described 7.
Statistical analysis
Results are expressed as mean±sem. Statistical analysis was performed using two-way repeated ANOVA. Subsequent post hoc analysis was performed using Bonferroni’s method. For all analyses, values of p<0.05 were assumed to be significant (n = number of donors (tracheae) whose cultured epithelial cells were used).
RESULTS
Effects of carbocisteine on rhinovirus infection in human tracheal epithelial cells
Exposing confluent human tracheal epithelial cell monolayers to RV2 (1×105 TCID50·mL-1) and RV14 (1×105 TCID50·mL-1) consistently led to infection. No detectable virus was revealed 1 h after infection. RV2 and RV14 were detected in culture medium at 6 h, and the viral content progressively increased during the period 6–24 h after infection (fig. 1a⇓ and c). Viral titres of supernatants collected 1–2 days after infection also contained significant levels of RV2 and RV14 (fig. 1a⇓ and c). Supernatant viral titres increased significantly with time for the first 48 h (p<0.05 using ANOVA).

Viral titres (50% tissue culture infective dose (TCID50) per millilitre) in supernatants of human tracheal epithelial cells obtained: a, c) at different times after exposure to 1×105 TCID50·mL-1 rhinovirus (RV)14 (a) and RV2 (c) in the presence (○) of carbocisteine (10 μM) or vehicle (PBS; •); and b, d) at different concentrations of carbocisteine 24–48 h after infection with RV14 (b) and RV2 (d). The cells were treated with carbocisteine or vehicle beginning 3 days before RV14 infection and continuing the end of the experiments. Data are presented as mean±sem (n = 5 tracheae). *: p<0.05 versus vehicle.
Treatment of the cells with carbocisteine significantly decreased the titres of RV2 and RV14 in supernatants 24 and 48 h after infection (fig. 1a⇑ and c). Furthermore, carbocisteine inhibited RV2 and RV14 infection in a concentration-dependent manner, the maximum effect being obtained at 10 and 30 μM (fig. 1b⇑ and d).
In order to determine whether or not RV14 infection or carbocisteine-induced cytotoxic effects on the cultured cells caused cell detachment from the tubes after the cells had formed a confluent sheet, cell numbers were counted after RV14 infection and after treatment with carbocisteine. Cell numbers were constant in the confluent epithelial cells in the control medium, and the coefficient of variation was small (7.3%; n = 15). Neither RV14 infection (1×105 TCID50·mL-1; 2 days) nor carbocisteine treatment (10 μM; 5 days) had any effect on cell number (data not shown). Cell viability, assessed by trypan blue exclusion 7, was consistently >96% in the carbocisteine-treated culture. RV14 infection and carbocisteine treatment (10 μM) did not alter the amount of LDH in the supernatants. The amount of LDH in the supernatants was 29±2 IU·L-1 before RV14 infection, 30±2 IU·L-1 2 days after RV14 infection (p>0.50; n = 5), and 30±2 IU·L-1 after carbocisteine treatment (10 μM; 5 days) (p>0.50; n = 5).
Effects of carbocisteine on viral RNA by PCR
No detectable RV14 was revealed before RV14 infection (data not shown). RV14 was detected 24 h after RV14 infection (fig. 2a⇓), and the amount of RV14 RNA in the cells was greater at 48 h than at 24 h (fig. 2b⇓). Carbocisteine (10 μM) caused a decrease in the amount of RV14 RNA in the cells 24 and 48 h after infection (fig. 2b⇓). The magnitude of the inhibitory effects of carbocisteine 48 h after infection was greater than that at 24 h (fig. 2b⇓).

a) Replication of rhinovirus (RV) 14 RNA (381 bp) from human tracheal epithelial cells 0, 24 and 48 h (lanes 1–3 and 4–6, respectively) after RV14 infection in the presence of 10 μM carbocisteine (lanes 4–6) or vehicle (PBS) alone (lanes 1–3) as detected by RT-PCR (β-actin RNA 217 bp). Data are representative of three different experiments. b) Corresponding data obtained by real-time quantitative RT-PCR (□: carbocisteine; ░: vehicle). A standard curve was constructed using the fluorescence emission signals and the cycle threshold by means of 10-fold dilutions of the total RNA, extracted from 1×105 of the 50% tissue culture infective dose (TCID50)·mL-1 RV14 in the supernatants of human embryonic fibroblasts 7 days after RV14 infection (1×104 TCID50·mL-1). Real-time quantitative RT-PCR for reduced glyceraldehye-3-phosphate dehydrogenase (GAPDH) was also performed using the same PCR products. RV RNA expression was normalised to the constitutive expression of GAPDH mRNA and expressed relative to cells treated with vehicle alone 24 h after infection. Data are presented as mean±sem (n = 5 tracheae). *: p<0.05; **: p<0.01 versus vehicle 24 h after infection; ##: p<0.01 versus vehicle 48 h after infection.
Effects of carbocisteine on susceptibility to rhinovirus 14 infection
Treatment of the cells with carbocisteine decreased their susceptibility to infection by RV14. The minimum dose of RV14 necessary to cause infection in cells treated with carbocisteine (10 μM; 3 days) (2.6±0.2 log TCID50·mL-1; p<0.05; n = 5) was significantly higher than that in cells treated with vehicle (PBS) (1.8±0.2 log TCID50·mL-1; n = 5).
Effects of carbocisteine on expression of intercellular adhesion molecule-1
Carbocisteine inhibited baseline ICAM-1 mRNA expression in the cells before RV14 infection (fig. 3a⇓). Carbocisteine reduced ICAM-1 mRNA expression by >50% compared with that of cells treated with vehicle (PBS) (fig. 3a⇓). Likewise, carbocisteine significantly reduced supernatant sICAM-1 concentrations before RV14 infection (fig. 3b⇓). In contrast, carbocisteine did not inhibit baseline LDL receptor mRNA expression in the cells before RV14 infection (fig. 3c⇓).

a) Expression of intercellular adhesion molecule (ICAM)-1 and low-density lipoprotein receptor (LDLR) mRNA in human tracheal epithelial cells 3 days after starting treatment with 10 μM carbocisteine (□) or PBS vehicle (░) detected by real-time quantitative RT-PCR. mRNA expression was normalised to the constitutive expression of reduced glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA. b) Soluble ICAM (sICAM)-1 concentration in cell supernatants 3 days after starting treatment with 10 μM carbocisteine or vehicle (0 μM carbocisteine). The medium was changed daily, and the supernatants collected 2–3 days after starting treatment. Results are mean±sem (n = 5 tracheae). ns: nonsignificant (versus control); *: p<0.05 versus control.
Effects of carbocisteine on cytokine production
Carbocisteine reduced the baseline secretion of IL-6 and IL-8 for 24 h before RV14 infection compared with that in the cells treated with vehicle (PBS) (table 1⇓). Furthermore, the secretion of IL-6 and IL-8 increased 24 h after RV14 infection. Carbocisteine also reduced the RV14 infection-induced secretion of IL-6 and IL-8 compared to that in the cells treated with vehicle 24 h after RV14 infection (table 1⇓).
Effects of carbocisteine on supernatant cytokine contents before and 24 h after rhinovirus(RV) 14 infection
Conversely, carbocisteine inhibited the baseline secretion of IL-1β for 24 h before RV14 infection compared with that in cells with no carbocisteine treatment (table 1⇑). In contrast, the secretion of IL-1β 24 h after RV14 infection did not differ from that before RV14 infection (table 1⇑). TNF-α was not detectable in supernatants for 24 h before and after RV14 infection.
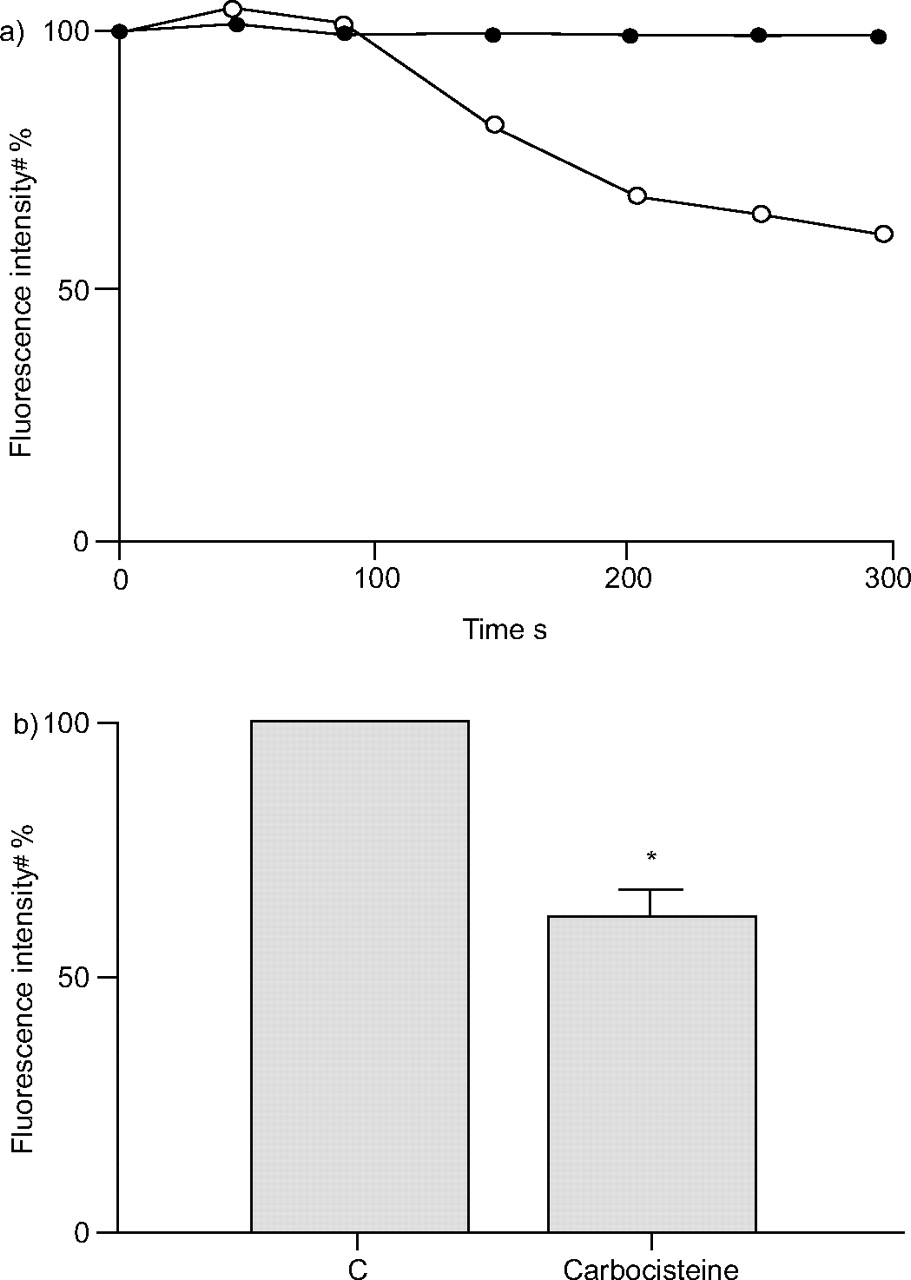
Effects of carbocisteine on the acidification of endosomes
Acidic endosomes in human tracheal epithelial cells were stained green using LysoSensor DND-189. Green fluorescence from acidic endosomes was observed in a granular pattern in the cytoplasm (data not shown), as previously described 7. Carbocisteine decreased the number and fluorescence intensity of acidic endosomes with green fluorescence in the cells over time (fig. 4a⇓). The fluorescence intensity of acidic endosomes in the epithelial cells treated with carbocisteine for 300 s was significantly reduced (fig. 4⇓).

a) Time course of changes in the intensity of green fluorescence from acidic endosomes in human tracheal epithelial cells after treatment with either 10 μM carbocisteine (○) or PBS vehicle (•). Inhibitors were administrated at time 0. b) Fluorescence intensity of acidic endosomes 300 s after the addition of 10 μM carbocisteine or vehicle (control (C)). The fluorescence intensity of acidic endosomes was measured in 100 human tracheal epithelial cells, and the mean expressed as a percentage of the control value (#). Data are presented as mean±sem (n = 5 tracheae). *: p<0.05 versus control.
Nuclear factor-κB DNA-binding activity in human tracheal epithelial cells
Baseline nuclear factor (NF)-κB DNA-binding activity was constant, and increased activation of NF-κB DNA-binding activity was present in the cells 120 min after RV14 infection (fig. 5⇓), as previously described 7. Carbocisteine reduced the increased activation of NF-κB occurring as a result of RV14 infection (fig. 5⇓).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Nuclear factor (NF)-κB DNA-binding activity of human tracheal epithelial cells before (lane 2) and 120 min after (lanes 3 and 4) rhinovirus (RV) 14 infection in the presence of 10 μM carbocisteine (lane 4) or PBS vehicle (lane 3) detected via electrophoretic mobility shift assay (lane 1: positive control (10 ng·mL-1 IL-1β plus 10 ng·mL-1 tumour necrosis factor-α for 24 h)). The cells were treated with carbocisteine or vehicle from 3 days before until 120 min after RV14 infection. Data are representative of three different experiments.
DISCUSSION
In the present study, it has been shown that a mucolytic drug, carbocisteine, reduced supernatant viral titres and viral RNA levels of a major group RV, RV14, in cultured human tracheal epithelial cells. Pretreatment with carbocisteine inhibited the expression of mRNA encoding ICAM-1, the receptor for the major group of RVs 14, as well as the supernatant concentrations of sICAM-1 before RV14 infection. Since the minimum dose of RV14 necessary to cause infection in cells treated with carbocisteine was significantly higher than that in cells treated with vehicle, carbocisteine may inhibit RV14 infection, at least partly, by reducing the production of its receptor, ICAM-1, as observed in human tracheal epithelial cells treated with dexamethasone 18 and erythromycin 7. Furthermore, carbocisteine reduced the fluorescence intensity of acidic endosomes in the epithelial cells. The magnitude of the inhibitory effects of carbocisteine on the fluorescence intensity of acidic endosomes was similar to that of bafilomycin A1 17 and erythromycin 7. Conversely, carbocisteine also reduced supernatant virus titres of a minor group RV, RV2, although it did not reduce the mRNA expression of the LDL receptor, the receptor for the minor group of RVs 24. Carbocisteine may also inhibit RV2 and RV14 RNA entry across acidic endosomes, as demonstrated in HeLa cells and human tracheal epithelial cells treated with bafilomycin A1 16, 17, 26 and erythromycin 18.
Various viruses have been reported to be responsible for exacerbations of disease in patients with COPD and bronchial asthma, including RVs, influenza virus and respiratory syncytial virus 2–4. Seemungal et al. 3 reported that 64% of COPD exacerbations were associated with a cold before the onset of exacerbations; 77 viruses were detected in 39% of COPD exacerbations, and 39 (58%) of these viruses were RVs. RVs are also associated with acute exacerbations in bronchial asthma 2. These findings suggest that RVs may be a major pathogen responsible for acute exacerbations of COPD and bronchial asthma.
Various mechanisms have been attributed to the pathogenesis of COPD and bronchial asthma exacerbations, including airway inflammation, airway oedema, bronchoconstriction and mucus hypersecretion 4. In addition, neutrophilic and eosinophilc inflammation in the exacerbations are associated with a variety of mediators, including IL-6 and IL-8, and the production and secretion of IL-6 and IL-8 are stimulated by RV14 in airway epithelial cells, as shown in the present study as well as previous studies 5–7. Furthermore, ICAM-1 interacts physiologically with leukocyte function-associated antigen-1, expressed on leukocytes, and thus plays a vital role in the recruitment and migration of immune effector cells to sites of local inflammation, as observed in patients with COPD 27. Therefore, reduced RV14 infection-induced production of IL-6 and IL-8 by carbocisteine, observed in the present study, may be associated with the modulation of airway inflammation after RV infection, and with the prevention of acute exacerbations of chronic obstructive bronchitis, as previously described 9.
In the present study, carbocisteine reduced the baseline production of cytokines, including IL-1β, IL-6 and IL-8, as well as ICAM-1, before RV14 infection. RV14 infection increased the production of IL-6 and IL-8, and carbocisteine also reduced RV14 infection-induced production of IL-6 and IL-8. The concentration of IL-1β in supernatants had not changed 24 h after RV14 infection, and TNF-α was not detectable in supernatants 24 h after RV14 infection, as previously reported 7, 18. Since carbocisteine reduced the RV14 titre in supernatants, the inhibitory effects of carbocisteine on RV14 infection, and on cytokine production itself, might be associated with the reduced production of IL-6 and IL-8 in the cells treated with carbocisteine after RV14 infection.
Increased activation of NF-κB was also apparent in cells 120 min after RV14 infection, as shown in previous studies 5, 7, and carbocisteine inhibited this activation. NF-κB increases the expression of genes encoding many cytokines, such as IL-6, IL-8 and ICAM-1 5. Therefore, a reduction in cytokine and ICAM-1 levels might be mediated via the carbocisteine-reduced activation of NF-κB.
Endosomal pH is thought to be regulated by vacuolar H+-ATPases 28 and ion transport across Na+/H+ antiporters 28. The inhibitors of Na+/H+ antiporters, 5-(N-ethyl-N-isopropyl)amiloride and N′′-[5-hydroxymethyl-3-(1H-pyrrol-1-yl)benzoyl]guanidine methanesulphonate (FR168888), as well as a vacuolar H+-ATPase inhibitor, bafilomycin, increase endosomal pH and inhibit RV14 infection in cultured human tracheal epithelial cells 7. Although there are no data to support this, the increased endosomal pH induced by carbocisteine in the present study may be associated with an inhibitory effect on vacuolar H+-ATPases or Na+/H+ antiporters in airway epithelial cells.
Recent reports revealed that the major group of RVs enters the cytoplasm of infected cells after binding to its receptor, ICAM-1 14. The entry of the RNA of a major group RV, RV14, into the cytoplasm of infected cells is thought to be mediated by destabilisation from receptor binding and endosomal acidification 15. The inhibitory effects of carbocisteine on infection by RV14 and its effects on endosomal pH in the present study are consistent with those of bafilomycin and erythromycin in previous studies 7, 16, 17. In addition, the inhibitory effects of carbocisteine on ICAM-1 expression in airway epithelial cells might also be associated with inhibitory effects on RV14 infection, as previously reported for the inhibitory effects of bafilomycin, erythromycin and dexamethasone 7, 17, 18.
In the present study, the inhibitory effects of carbocisteine were observed over a small range of concentrations, and there is no complete inhibition of infection. The precise reason for this is uncertain. However, the magnitude of the inhibitory effects of carbocisteine on supernatant RV titres was smaller than those of dexamethasone and erythromycin 7, 18. The weak inhibitory effects of carbocisteine might show a small range of response and incomplete inhibition of infection. The magnitude of the inhibitory effects of carbocisteine on supernatant sICAM-1 concentrations was smaller than those of dexamethasone and erythromycin on ICAM-1 protein expression 7, 18, although the magnitude of the inhibitory effects of carbocisteine on the fluorescence intensity of acidic endosomes was similar to those of bafilomycin A1 17 and erythromycin 7. Therefore, smaller inhibitory effects of carbocisteine on ICAM-1 expression might be partly associated with lesser inhibition by carbocisteine of RV infection compared with the inhibitory effects of dexamethasone and erythromycin.
Carbocysteine exerts an anti-oxidant action by scavenging reactive oxygen intermediates 29. Indeed, SCMC-Lys reduces the concentration of 8-isoprostane, a prostaglandin-like compound, during peroxidation of membrane phospholipids by reactive oxygen species in the breath condensate of acute COPD patients 10. Furthermore, bafilomycin A1, a blocker of vacuolar H+-ATPase, reduces endosome acidification 17 in airway epithelial cells and reduces production of reactive oxygen species in alveolar macrophages 30, suggesting a relationship between antioxidant effects and the reduced acidification of endosomes. Reduced production of inflammatory cytokines and ICAM-1 by carbocisteine, in the present study, also suggests anti-inflammatory effects of carbocisteine, as shown in previous studies that demonstrated reduced IL-6 concentrations in breath condensate caused by SCMC-Lys in COPD patients 10 and reduced numbers of inflammatory cells in airways in rats caused by SCMC-Lys 11 and SCMC 12.
In summary, this is the first report that a mucolytic drug, carbocisteine, inhibits infection by rhinovirus 14 and decreases the susceptibility of cultured human tracheal epithelial cells to rhinovirus 14 infection, probably through the inhibition of intercellular adhesion molecule-1 expression and endosomal acidification. Carbocisteine also inhibited rhinovirus 2 infection, probably through the inhibition of endosomal acidification. Carbocisteine reduced baseline and rhinovirus infection-induced release of pro-inflammatory cytokines, such as interleukin-6 and -8, in supernatants. Carbocisteine may inhibit infection by the major and minor group of rhinoviruses, and modulate inflammatory responses in the airway epithelial cells after rhinovirus infection.
Acknowledgments
The authors would like to thank G. Crittenden for reading the manuscript.
- Received May 18, 2005.
- Accepted February 17, 2006.
- © ERS Journals Ltd
References










