





Abstract
The antioxidant N-acetylcysteine (NAC) prevented sepsis-induced diaphragmatic dysfunction. As an indirect antioxidant NAC was shown to induce superoxide dismutase (SOD) activity in immune cells from endotoxaemic mice. The aim of this study was to assess whether NAC acts as an indirect antioxidant by inducing manganese (Mn)-SOD activity in the diaphragms of endotoxaemic rats, while preventing muscle dysfunction.
A controlled study was conducted, in which protein carbonylation, Mn-SOD, catalase, and 3-nitrotyrosine immunoreactivity were detected using immunoblotting and immunohistochemistry in rat diaphragms. Six groups were studied for 24 h after a saline (control) or lipopolysaccharide (LPS; 20 mg·kg−1) i.p. injection in the absence and presence of NAC pre-treatment (either 1.5 or 3 mmol·kg−1·24 h−1 for 7 days, oral administration). Diaphragm mitochondrial Mn-SOD activity and respiratory muscle function were also determined.
Within 24 h, LPS induced maximal inspiratory pressure reduction, increasing diaphragmatic protein carbonylation and nitration. Pre-treatment with 3 mmol·kg−1 NAC clearly increased muscle Mn-SOD protein content and activity in both LPS- and saline-injected animals, while reducing protein carbonylation and nitration, and partially preventing the LPS-induced respiratory muscle dysfunction.
Data produced from this study indicate that high doses of N-acetylcysteine induces manganese superoxide dismutase, as well as preserves its activity, possibly by preventing nitration of critical tyrosine residues of the enzyme.
- Antioxidant enzymes
- endotoxaemia
- oxidative stress
- protein tyrosine nitration
- respiratory muscle dysfunction
It has been well established that the ventilatory muscle dysfunction observed in sepsis is attributable to the contribution of different factors such as excessive production of pro-inflammatory cytokines, reactive oxygen species (ROS), nitric oxide, and bacterial lipopolysaccharide (LPS). LPS is the major component of the outer membrane of Gram-negative bacteria, playing a crucial role in triggering the inflammatory cascade and the haemodynamic events in sepsis.
Over the last decade, accumulating evidence has shown that oxidative stress, defined as the imbalance between oxidants and antioxidants in favour of the former, is involved in the sepsis-induced muscle contractile dysfunction 1–4. Recent observations by Barreiro et al. 5 have indicated that protein tyrosine nitration, the most commonly studied covalent modification of proteins attributed to nitric oxide, was also increased in the diaphragms of endotoxaemic rats. The view has also been put forward that haemoxygenases 6, 7 have protective effects, probably by reducing muscle oxidative stress levels in sepsis-induced diaphragmatic dysfunction. In another study, mechanical ventilation, through reduction of both oxidative and mechanical stress, was also shown to prevent the diaphragmatic dysfunction induced by LPS administration 8.
N-acetylcysteine (NAC) is a molecule with antioxidant properties, as it can react either directly or indirectly with a variety of ROS. For instance, NAC reduced the rate of development of diaphragm fatigue during repetitive contractions 9 and depressed in vitro contractility of nonfatigued muscles in a dose-dependent manner 10 through direct reduction of ROS-mediated damage to either sarcoplasmic reticulum or mitochondrial membranes 9, 10. As an indirect antioxidant, NAC was shown to induce superoxide dismutase (SOD) activity in immune cells from mice with lethal endotoxic shock previously treated with NAC, while oxidative stress indices decreased significantly 11, 12. Based on this information, the authors hypothesised whether NAC might act as an indirect antioxidant via the stimulation of primary intracellular antioxidant defences within the muscle fibres in a model of endotoxaemia. Therefore, the study was aimed at determining whether the content and activity of manganese (Mn)-SOD, an essential enzyme to protect against oxidant injury, were enhanced by NAC pre-treatment in the diaphragm muscles of endotoxaemic rats, thereby preventing muscle dysfunction.
MATERIALS AND METHODS
Animals
Experimental groups
Six groups of pathogen-free male Sprague-Dawley rats (275–300 g, n = 8 per group) of identical age (8 weeks) were studied 24 h after LPS or saline (control groups) intraperitoneal injection in the absence and presence of NAC pre-treatment. The distribution of the animal groups was as follows: groups one and two were given an intraperitoneal injection of either 20 mg·kg−1 of Escherichia coli LPS (serotype 055: B5; Sigma-Aldrich, Schnelldorf, Germany) or normal saline; groups three and four were pre-treated with 1.5 mmol·kg−1 and 3 mmol·kg−1 NAC, respectively, which was administered orally, using a 14-mm gauge needle every 24 h for 7 consecutive days 13 before the LPS injection; groups five and six were also pre-treated with both 1.5 mmol·kg−1 and 3 mmol·kg−1 NAC, respectively, every 24 h for 7 consecutive days before the saline injection. Although the mortality ratio among the groups was very low (∼8%), those animals which died during the course of the experimental protocol were immediately discarded and always replaced with more rats who again underwent the different procedures. All animals were anaesthetised with penthothal (100 mg·kg−1; B. Brown Medical S. A., Barcelona, Spain), and were killed 24 h after the injection. The diaphragm was then quickly excised and either frozen in liquid nitrogen or embedded in paraffin for immunohistochemical use.
Study design
This controlled study was designed in accordance with both the ethical standards on animal experimentation in the IMIM (Barcelona, Spain), Basque Country University (Bizkaia, Spain) and McGill University (Montreal, Quebec, Canada), as well as the Helsinki convention for the use and care of animals. All experiments were approved by the Animal Research Committees for the respective institutions.
Physiological studies
Inspiratory pressure measurements
The animals were first placed, fully awake, in a polyurethane plethysmographic chamber, the anterior part of which contained a two-way valve (model 2200; Hans Rudolph, Kansas City, MO, USA). Both the neck and head of each rat were held by a latex membrane, positioned in close proximity to the valve, while a piston placed in the posterior part of the chamber pushed the animal forward to keep it immobilised. A plastic tube attached to the inspiratory branch of the valve and connected to a pressure transducer (ML 140; ADI Instruments, Castle Hill, Australia) was used to measure inspiratory pressure in all rats. Maximal inspiratory pressure (PI,max) was determined by total occlusion of the inspiratory circuit of the Hans Rudolph valve for 30 s, and this manoeuvre was performed a minimum of two times, separated by a time period of <60 s 15. PI,max was the highest value obtained from the two different measurements. In a group of rats pre-treated with 3 mmol·kg−1 of NAC (n = 8), PI,max was measured at three different time-points: 1) at baseline (after a 7-day acclimatisation period), 2) after the 7-day pre-treatment with 3 mmol·kg−1 NAC, and 3) 24 h after either the LPS or the saline i.p. injection (before the pentothal administration and animal sacrifice). In another group of animals not pre-treated with NAC (LPS-control and saline-control groups, n = 8 each), PI,max was measured: 1) at baseline and 2) 24 h after either the LPS or the saline i.p. injection.
Biological muscle studies
Immunoblotting
The levels of oxidative and nitrosative stress were assessed, as extensively described elsewhere 16–18. Accordingly, total carbonyl groups, which were detected through derivatisation to 2,4-dinitrophenylhydrazone (DNP) 19, Mn-SOD, catalase, and protein tyrosine nitration (3-nitrotyrosine immunoreactivity), were determined using: anti-DNP moiety (Oxyblot kit; Chemicon International Inc., Temecula, CA, USA) 18; anti-Mn SOD (StressGen, Victoria, BC, Canada), anti-catalase antibody (Calbiochem, San Diego, CA, USA); and anti-3-nitrotyrosine (Cayman Chemical Inc., Ann Arbor, MI, USA) antibodies.
Immunohistochemistry
Immunohistochemical analysis was performed as described elsewhere 16, 18. All sections (3 μm) were deparaffinated, and incubated with anti-DNP moiety, anti-Mn-SOD, anti-catalase and anti-3-nitrotyrosine primary antibodies, followed by incubation with biotinylated secondary antibodies, and with both horseradish peroxidase-conjugated streptavidin and diaminobenzidine (Dako Corporation, Carpinteria, CA, USA).
Isolation of muscle mitochondria
The specific activity of Mn-SOD was determined by isolating mitochondria from the rat diaphragms as described elsewhere 5, 20.
Superoxide dismutase assay
A commercially available superoxide dismutase assay kit (Calbiochem, San Diego, CA, USA) was used according to the manufacturer's instructions to selectively determine the activity of the mitochondrial isoform, following the entire procedure described by Nebot et al. 21. Samples were always run in triplicate and their corresponding activity was expressed as the mean value of the three measurements.
Statistical analysis
Data are presented as mean±sd. ANOVA, Tukey corrected, for multiple comparisons, was used to compare data obtained within the different groups; p≤0.05 was considered significant.
RESULTS
Functional assessment
Within 24 h, LPS injection elicited a significant decrease in PI,max (43%) compared with baseline values (fig. 1⇓). Furthermore, only in those rats pre-treated with 3 mmol·kg−1 NAC, was the decline in PI,max partially and significantly attenuated (30%) with respect to baseline (fig. 1⇓). PI,max values were not significantly modified after the 7-day pre-treatment period with either 3 mmol·kg−1 or 1.5 mmol·kg−1 NAC only (data not shown).

The significant fall (43%) from baseline (maximal inspiratory pressure: PI,max) average value of all the animals after a 7-day acclimatisation period, n = 16, represented as the 100%) in PI,max 24 h after lipopolysaccharide (LPS) injection (average PI,max measurements in those animals only injected with LPS in the absence of N-acetylcysteine (NAC) pre-treatment, n = 8) was partially and significantly attenuated (30%) by previous treatment with 3 mmol·kg−1 NAC/24 h for 7 days (average of PI,max measurements 24 h after the LPS injection in those rats pre-treated with 3 mmol·kg−1 NAC, n = 8). PI,max was determined by total occlusion of the inspiratory circuit of the Hans Rudolph valve. Animals were always fully awake while this manoeuvre was being performed. *: p<0.05; ***: p<0.001.
Protein carbonylation
Anti-DNP antibody detected different positive protein bands in the diaphragms of both endotoxaemic animals and controls (fig. 2⇓). Within 24 h, LPS induced a significant rise in protein carbonylation in the rat diaphragms compared with animals injected with saline (fig. 3⇓). Diaphragms from animals pre-treated with both 3 mmol·kg−1 and 1.5 mmol·kg−1 NAC showed a significant reduction in carbonyl group formation 24 h after either LPS or saline injection (fig. 3⇓). Immunostaining with anti-DNP antibody revealed the presence of abundant carbonyl groups diffusely localised within the diaphragm fibres 24 h after LPS injection (fig. 4⇓). This intensity was noticeably reduced when animals were pre-treated with 3 mmol·kg−1 NAC (fig. 4⇓).

Representative examples of diaphragm carbonyl groups 24 h after either lipopolysaccharide (LPS) or saline injection of rats untreated and treated with either 1.5 mmol·kg−1 or 3 mmol·kg−1 N-acetylcysteine (NAC). Several carbonylated proteins were detected. mw kD: molecular weight in kilodaltons; Ctl: control.
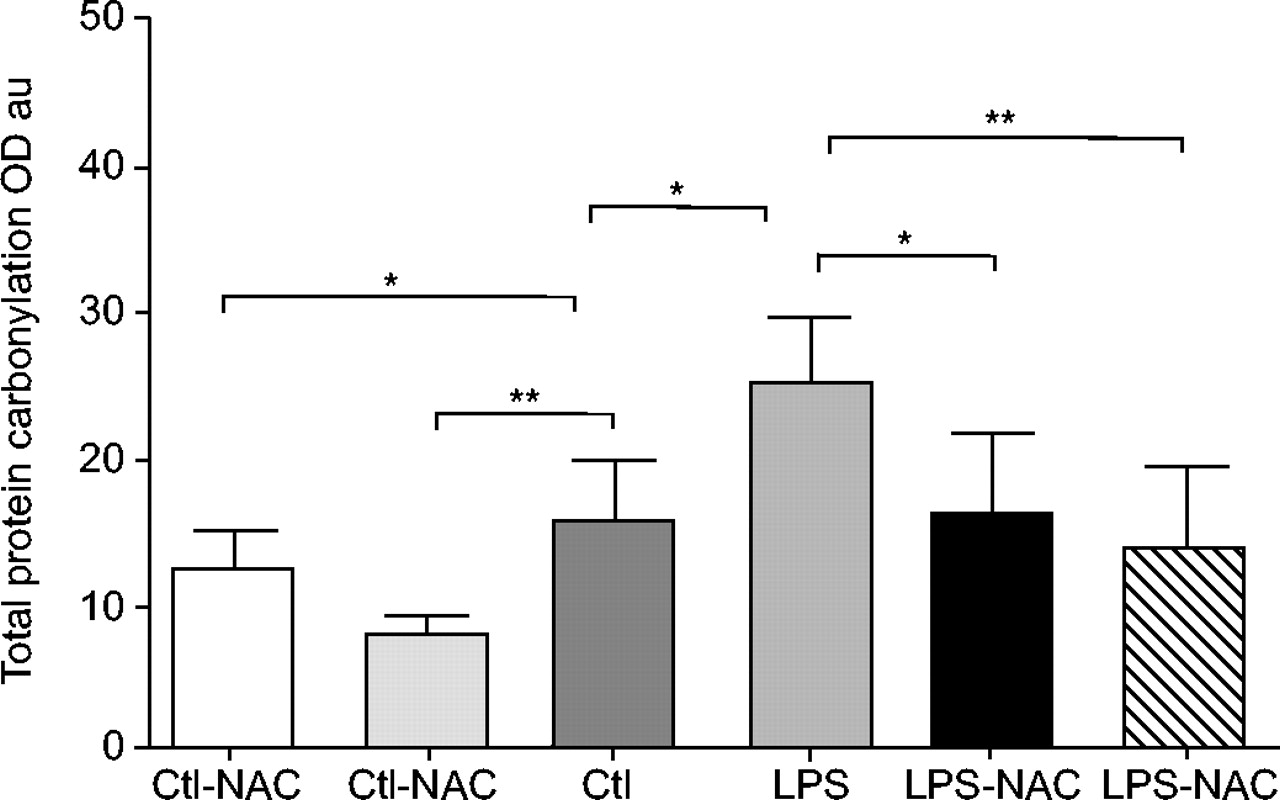
Mean values±sd of total protein carbonylation were higher in the diaphragms of lipopolysaccharide (LPS)-injected animals compared with saline-injected rats. Previous treatment with either dose of N-acetylcysteine (NAC) significantly reduced protein carbonyl content 24 h after either LPS or saline injection. Note that a 3 mmol·kg−1 dose induced a greater decrease in protein carbonylation in LPS-injected animals. OD: optical density; au: arbitrary units; Ctl: control; *: p<0.05; **: p<0.01.

Immunohistochemical localisation of carbonyl-modified proteins in diaphragm fibres of one lipopolysaccharide-injected animal (a) and another pre-treated with 3 mmol·kg−1 N-acetylcysteine (b). Anti-dinitrophenylhydrazone (DNP) antibody detected positive staining diffusely localised within the muscle fibres (a and b). Avoiding the derivatisation process eliminates positive-protein carbonylation staining (negative controls: c and d). Furthermore, replacement of the primary anti-DNP antibody with secondary antibodies completely eliminated positive carbonyl-modified protein staining (negative controls: e and f). Scale bars = 50 μm.
Antioxidant enzymes
The enzyme Mn-SOD was present in all rat diaphragms (fig. 5⇓). Within 24 h, LPS induced a significant rise in diaphragmatic Mn-SOD protein content compared with animals injected with saline (fig. 6⇓). Diaphragms from animals pre-treated with 3 mmol·kg−1 NAC showed a significant increase in Mn-SOD protein content 24 h after either LPS or saline injection (fig. 6⇓). However, in those animals pre-treated with 1.5 mmol·kg−1 NAC, Mn-SOD content was only significantly increased 24 h after LPS injection and not after saline injection (fig. 6⇓). Within 24 h, LPS injection induced positive staining of Mn-SOD diffusely localised in the diaphragm fibres (fig. 7⇓), and this intensity was markedly enhanced in animals that had been pre-treated with 3 mmol·kg−1 NAC (fig. 7⇓). Within 24 h, LPS did not induce any significant difference in diaphragmatic Mn-SOD activity compared with animals injected with saline (fig. 8⇓). Diaphragms from animals pre-treated with 3 mmol·kg−1 NAC showed a significant increase in Mn-SOD activity 24 h after either LPS or saline injection (fig. 8⇓).

Representative examples of protein expression of manganese superoxide dismutase (Mn-SOD) in the diaphragms of both lipopolysaccharides (LPS)- and saline-injected rats, untreated and treated with either 1.5 mmol·kg−1 or 3 mmol·kg−1 N-acetylcysteine (NAC). Corresponding positive control (rat brain mitochondria) is indicated accordingly. mw kD: molecular weight in kilodaltons: base pairs; Ctl: control.
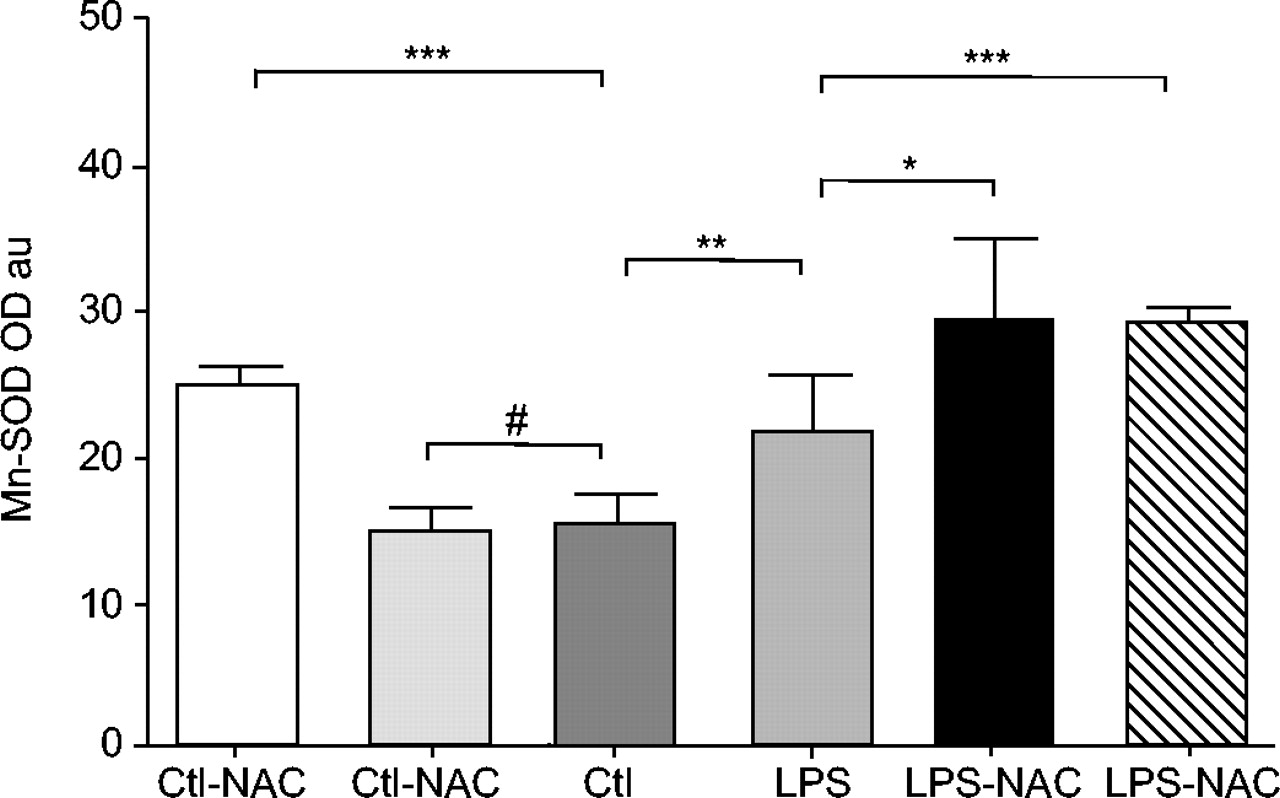
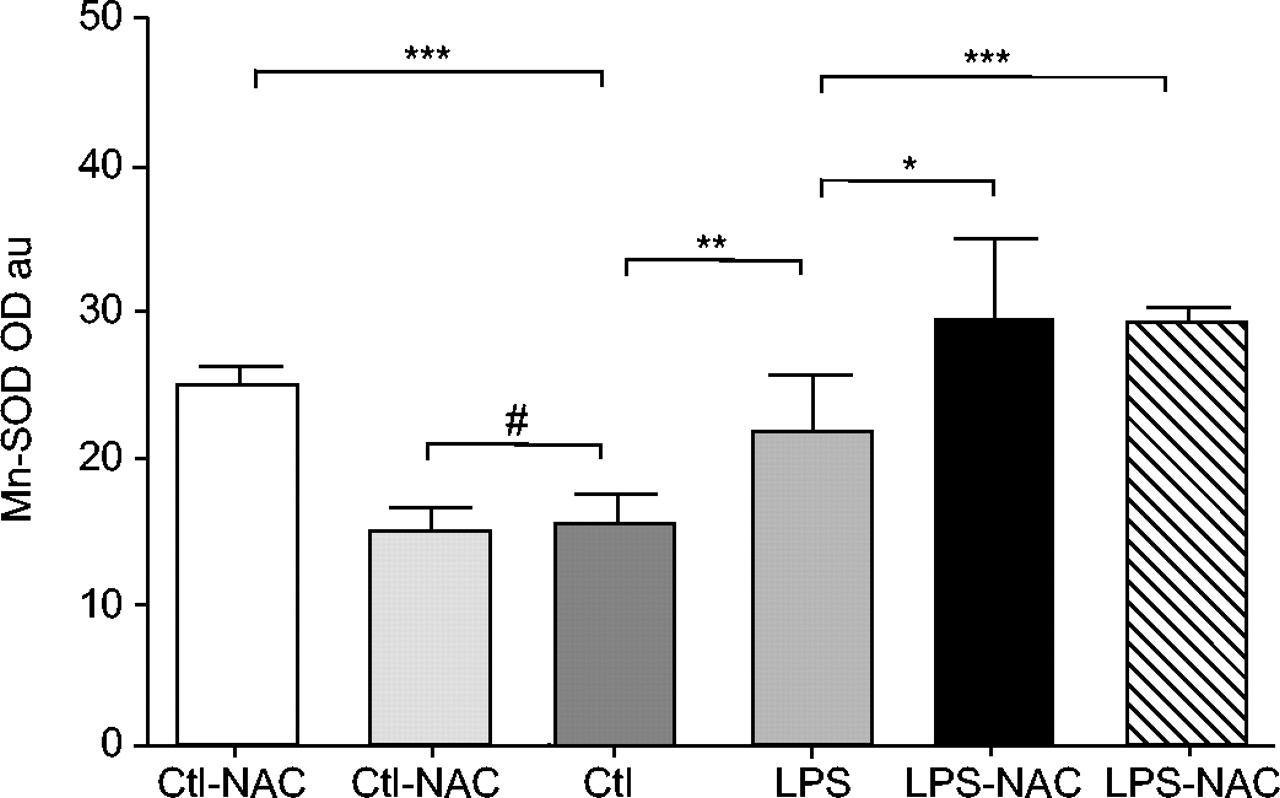
Mean±sd values of manganese superoxide dismutase (Mn-SOD) were increased in rat diaphragms 24 h after lipopolysaccharide (LPS) injections, as compared with muscles of saline-injected animals. Previous treatment with 3 mmol·kg−1 N-acetylcysteine (NAC) significantly increased diaphragm Mn-SOD protein content in both LPS- and saline-injected rats. Note that a 1.5 mmol·kg−1 dose also induced an increase in muscle Mn-SOD protein content only in LPS-injected animals. Ctl: control. #: nonsignificant. *: p<0.05; **: p<0.01; ***: p<0.001.

Immunohistochemical localisation of manganese superoxide dismutase (Mn-SOD) within the muscle fibres of lipopolysaccharide (LPS)-injected animals either untreated (a) or treated (b) with 3 mmol·kg−1 N-acetylcysteine (NAC). Note that Mn-SOD staining was more intense in the fibres of animals pre-treated with 3 mmol·kg−1 NAC. Replacement of the primary anti-Mn-SOD antibody with nonspecific secondary antibodies completely eliminated positive staining (negative controls: c and d). Scale bars = 50 μm.
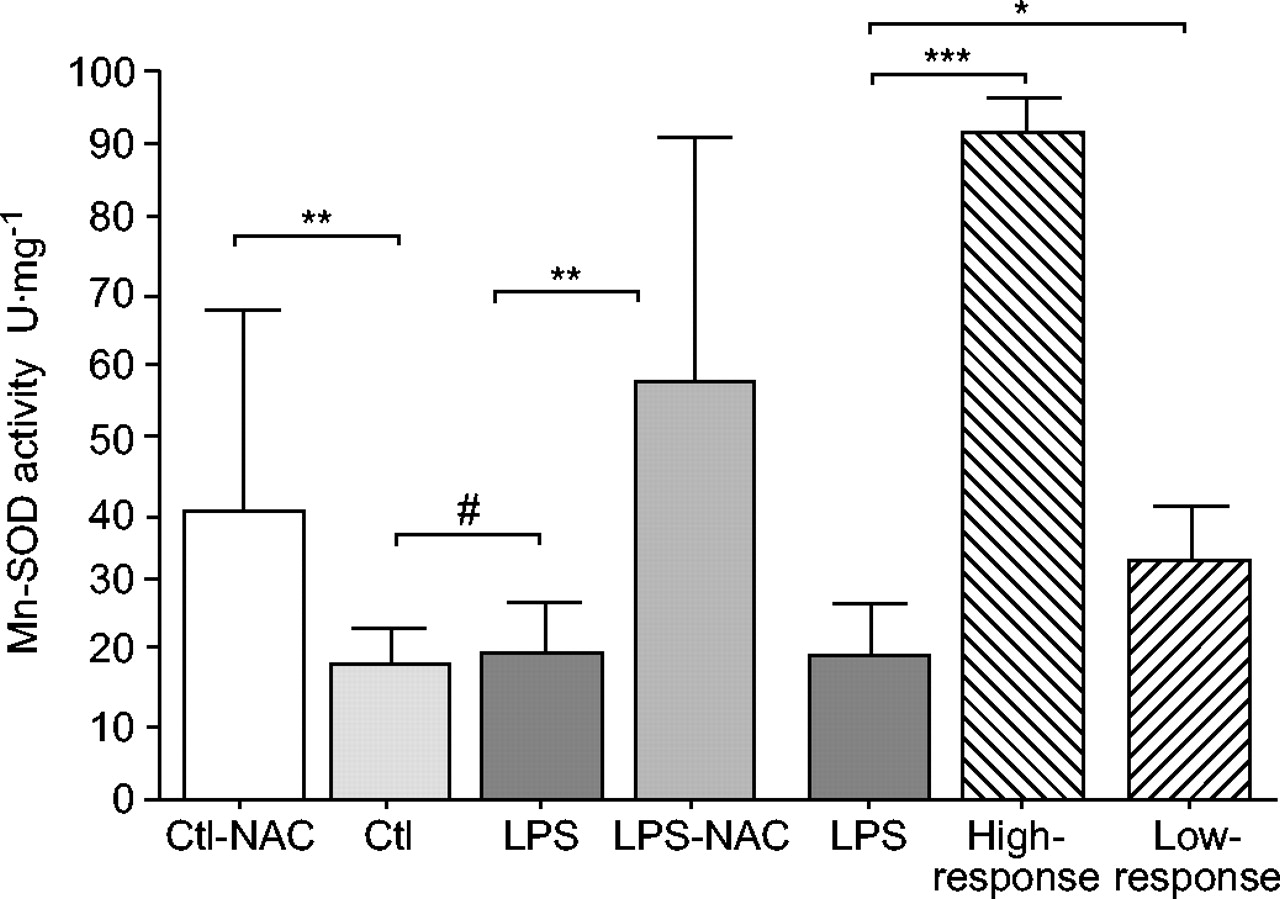
Animals pre-treated with 3 mmol·kg−1 N-acetylcysteine (NAC) showed increased diaphragm activity of the mitochondrial enzyme manganese superoxide dismutase (Mn-SOD) in either lipopolysaccharide (LPS)- or saline-injected rats. Note that in those animals not treated with NAC muscle Mn-SOD activity did not increase during the 24 h following LPS injection with respect to saline-injected rats. Ctl: control. *: p<0.05; **: p<0.01; ***: p<0.001; #: nonsignificant.
The enzyme catalase was present in all rat diaphragms (fig. 9⇓). Within 24 h, LPS induced no significant difference in diaphragmatic catalase protein content compared with animals injected with saline (fig. 10⇓). Diaphragms from animals pre-treated with either 3 mmol·kg−1 or 1.5 mmol·kg−1 NAC showed no significant difference in catalase content 24 h after either LPS or saline injection (fig. 10⇓). Within 24 h, LPS injection induced positive staining of catalase within the cytosol of diaphragm fibres in the absence and presence of 3 mmol·kg−1 NAC treatment (fig. 11⇓).

Representative examples of protein expression of catalase in the diaphragms of lipopolysaccharides (LPS)- and saline-injected rats, untreated and treated with either 1.5 mmol·kg−1 or 3 mmol·kg−1 N-acetylcysteine (NAC). mw kD: molecular weight in kilodaltons: base pairs +C: positive control (rat erythrocytes); Ctl: control.

Mean±sd values of muscle protein catalase content in both lipopolysaccharides (LPS)- and saline-injected animals, untreated and treated with 1.5 mmol·kg−1 and 3 mmol·kg−1 N-acetylcysteine (NAC). Diaphragmatic levels of catalase did not differ among the different groups of animals. Ctl: control. #: nonsignificant.

Immunohistochemical localisation of catalase in the cytosol of the diaphragm fibres of lipopolysaccharides-injected animals untreated (a) and treated (b) with 3 mmol·kg−1 N-acetylcysteine. Replacement of the primary anticatalase antibody with nonspecific secondary antibodies completely eliminated positive catalase staining (negative controls: c and d). Scale bars = 50 μm.
Protein tyrosine nitration
Several tyrosine-nitrated protein bands were detected in all muscles (fig. 12⇓). Within 24 h, LPS induced a significant rise in diaphragmatic 3-nitrotyrosine immunoreactivity compared with animals injected with saline (fig. 13⇓). Diaphragms from animals pre-treated with 3 mmol·kg−1 NAC showed a significant decrease in protein tyrosine nitration 24 h after either LPS or saline injection (fig. 13⇓). However, in those animals pre-treated with 1.5 mmol·kg−1 NAC, 3-nitrotyrosine immunoreactivity was only slightly, nonsignificantly, reduced 24 h after LPS injection (fig. 13⇓). Within 24 h, LPS injection induced positive staining of 3-nitrotyrosine diffusely localised in the diaphragm fibres (fig. 14⇓), and this intensity was clearly reduced when animals were pre-treated with 3 mmol·kg−1 NAC (fig. 14⇓).

Representative examples of diaphragm protein tyrosine nitration (total 3-nitrotyrosine immunoreactivity) 24 h after both lipopolysaccharides (LPS) and saline injection in rats, untreated and treated with either 1.5 mmol·kg−1 or 3 mmol·kg−1 N-acetylcysteine (NAC). Several tyrosine-nitrated proteins were detected. Ctl: control; mw KD: molecular weight in kilodaltons.

Mean values±sd of total diaphragm 3-nitrotyrosine immunoreactivity were higher 24 h after lipopolysaccharides (LPS) injection compared with those of saline-injected animals. Pre-treatment with 3 mmol·kg−1 N-acetylcysteine (NAC), but not with 1.5 mmol·kg−1 NAC, significantly reduced muscle protein tyrosine nitration in both LPS- and saline-injected rats. Ctl: control. *: p<0.05; **: p<0.01; ***: p<0.001; #: nonsignificant.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Immunohistochemical localisation of diaphragm 3-nitrotyrosine 24 h after lipopolysaccharide (LPS) injection in both untreated (a) and 3 mmol·kg−1 NAC pre-treated (b) animals. Anti-3-nitrotyrosine antibody detected positive staining diffusely localised within the muscle fibres (a and b). Note that the staining was less intense in the fibres of LPS-injected animals pre-treated with 3 mmol·kg−1 N-acetylcysteine (NAC). Replacement of the primary anti-3-nitrotyrosine antibody with nonspecific secondary antibodies completely eliminated positive staining (negative controls: c and d). Scale bars = 50 μm.
DISCUSSION
The main findings of the present study are: 1) 24 h after LPS injection, PI,max was significantly reduced while both diaphragm protein carbonylation and nitration as well as Mn-SOD protein content were increased; 2) pre-treatment of rats with 3 mmol·kg−1 NAC partially and significantly attenuated the fall in PI,max, while reducing muscle protein carbonylation and nitration levels in both LPS- and saline-injected animals; and 3) pre-treatment with 3 mmol·kg−1 NAC resulted in an increase in muscle Mn-SOD activity in both LPS- and saline-injected rats.
Oxidative stress and endotoxaemia-induced respiratory muscle dysfunction
As previously shown 22–25, the current study confirms that within 24 h LPS induced a reduction in the respiratory muscle force of endotoxaemic animals, and an increase in diaphragmatic both protein carbonylation and nitration levels 1–8, 26. The administration of scavengers of a variety of free radicals, as well as inhibitors of nitric oxide synthases, was shown to prevent sepsis-induced ventilatory muscle dysfunction in animals 1, 3, 26, leading to the conclusion that various free-radical species are likely to participate in that respiratory muscle dysfunction 26. In line with this, recent evidence has shown that the absence of haem oxygenase activity within the diaphragm fibres increased the levels of oxidative stress and significantly reduced the ability of those fibres to generate force 6, 7. On this basis, it could be argued that the major species displaying the greatest oxidative modifications in those diaphragms are most likely to be proteins directly involved in muscle function. Indeed, in a model of endotoxaemic rats, Callahan et al. 4 demonstrated that oxidants altered muscle contractile protein force-generating capacity of diaphragm fibres. More recently, Barrerio et al. 27 have also gained some insight into specific physiological and metabolic processes that are targeted by ROS within the diaphragm fibres in endotoxaemic rats. Specifically, protein carbonylation, was predominant in those proteins involved in either glycolysis, adenosine triphosphate (ATP) production, or myofibrillar contraction, and the activity of these enzymes was inversely related to their protein carbonylation levels 27.
Effects of N-acetylcysteine on lipopolysaccharide-induced respiratory muscle dysfunction
In the present study, pre-treatment with 3 mmol·kg−1 NAC conferred protection against endotoxaemia by both partially reversing the respiratory muscle dysfunction and by diminishing indirect indices of ROS production in the rat diaphragms. NAC and other antioxidants influence muscle-fibre function by scavenging ROS. The exact molecular mechanisms whereby oxidant removal may affect muscle cell function are not yet fully understood. NAC could act as a direct antioxidant in skeletal muscles fibres by protecting different molecular sites such as sarcoplasmic reticulum, sarcolemma and/or mitochondria proteins from oxidative damage in fatiguing muscles and/or endotoxaemia 1, 9, 28, 29. Indeed, NAC was shown to have beneficial antioxidant effects on muscles during repetitive isometric contractions in both experimental animal 9, 10 and human studies 30, in quadriceps muscles of patients with severe chronic obstructive pulmonary disease 31, in endotoxaemic rats 1, and in the diaphragmatic dysfunction of streptozotocin-induced diabetic rats 14.
Mechanisms of manganese superoxide dismutase induction and its role as an antioxidant
In the present study, pre-treatment of rats with 3 mmol·kg−1 NAC, rather than with the lower dose, induced an increase in both Mn-SOD protein content and activity in the diaphragms of either LPS- or saline-injected rats. SOD is present in all aerobic organisms, and specifically Mn-SOD appears to be essential for life 32. Mitochondria are a major source of ROS production in muscle fibres and mitochondrial integrity is preserved by the scavenging of superoxide anion by Mn-SOD 33. Mn-SOD protein expression is induced by different stimuli including LPS 34, as also shown in the current study, but experiments demonstrating only Mn-SOD mRNA induction should be viewed with caution, as they do not necessarily correlate with the enzyme protein or activity levels.
MacMillan-Crow et al. 35–37 have recently demonstrated that nitration of three of the nine tyrosine residues of the Mn-SOD enzyme is mediated by petroxynitrite. The biological importance of this phenomenon is the inhibition of the enzyme activity in the models in question 35–37 through nitration of critical tyrosine residues 33. On this basis, it appears that Mn-SOD induction clearly needs to be analysed in terms of protein expression and especially activity, since gene upregulation of the enzyme will not correlate with equal increases in the enzyme content and/or activity in those conditions known to produce increased peroxynitrite formation, such as endotoxaemia 5. Hence, it can be concluded from the current findings that NAC probably acts as an indirect antioxidant, increasing Mn-SOD protein content and activity in the endotoxaemic diaphragms by scavenging peroxynitrite, at least at the mitochondrial level. In line with this, it has already been demonstrated 5 that LPS-mediated increase in protein tyrosine nitration was limited to the mitochondria and membrane compartments of septic rat diaphragms. Moreover, such a conclusion is supported by the fact that in this study, both protein tyrosine nitration and carbonylation levels were significantly reduced in the diaphragms of the endotoxaemic rats pre-treated with 3 mmol·kg−1 NAC. Other studies where increased SOD activity and superoxide anion reduction were shown in peripheral blood cells of endotoxaemic rats pre-treated with NAC 11, 12 also hold true for this hypothesis. Finally, the induction of Mn-SOD protein content and activity in nonseptic (control) muscles of animals pre-treated with 3 mmol·kg−1 NAC can also be explained in terms of a peroxynitrite-scavenging mechanism as implied by the lower levels of both protein carbonylation and nitration detected in those diaphragms.
The molecular targets whereby NAC might scavenge peroxynitrite and eventually prevent nitration of tyrosine residues in Mn-SOD enzyme of diaphragms remain unknown. However, it could be proposed that the redox-sensitive nuclear factor-κB might induce a decrease in both ROS and nitric oxide production within those muscle fibres, as shown in other models and diseases 38. Clearly, future studies are required to fully elucidate the cellular and molecular sites whereby NAC induces Mn-SOD activity as well as to explore the tyrosine nitration profile of this enzyme and precise functional implications in endotoxaemic muscles.
Finally, it is worth mentioning that the effects on muscles induced by pre-treatment with 1.5 mmol·kg−1 NAC differ from those achieved by the higher dose of this compound. For instance, diaphragmatic protein nitration levels were not significantly modified 24 h after either LPS or saline injection in those animals pre-treated with the lower dose. In addition, muscle Mn-SOD content was not increased in the saline-injected animals, and Mn-SOD activity was not modified for up to 24 h after LPS or saline injection (data not shown) when animals were pre-treated with 1.5 mmol·kg−1 NAC. From these findings, the authors conclude that in this experimental model of endotoxaemia, NAC reduces muscle oxidative stress levels and reverses contractility, but not in a dose-dependent fashion. The hypothesis of the existence of a peroxynitrite-mediated mechanism of Mn-SOD inactivation that could be reversed by NAC treatment would also fit here, since it is very likely that a “threshold-like” dose of the antioxidant NAC is required to trigger those events.
In summary, the current study proposes an indirect mechanism of a high dose of N-acetylcysteine as an antioxidant in endotoxaemic muscles, via manganese superoxide dismutase induction, by preventing peroxynitrite-mediated nitration of tyrosine residues in the active site of the antioxidant enzyme. Further studies are needed to explore the clinical and therapeutic applicability of these effects and mechanisms in septic patients.
Acknowledgments
The authors would like to thank B. de la Puente, A. Llorens, F. Sánchez, and J. Sellarés for their technical assistance in the laboratory, P. Peretti for his technical support in the preparation of the figures, and R. Marshall for his aid in editing the manuscript (URMAR, IMIM-UPF, Barcelona, Spain).
- Received January 11, 2005.
- Accepted August 21, 2005.
- © ERS Journals Ltd
References









