





Abstract
Thiazolidinedione rosiglitazone and 15-deoxy-Δ12,14-prostaglandin J2 (15d-PGJ2), are two peroxisome proliferator-activated receptor (PPAR)-γ ligands. The aim of this study was to investigate the effect of rosiglitazone and 15d-PGJ2 on the lung injury caused by bleomycin administration.
Mice subjected to intratracheal administration of bleomycin developed significant lung injury. An increase in immunoreactivity to nitrotyrosine, poly(ADP ribose) polymerase (PARP) and inducible nitric oxide synthase as well as a significant loss of body weight and mortality was observed in the lung of bleomycin-treated mice.
Administration of the two PPAR-γ agonists rosiglitazone (10 mg·kg−1 i.p.) and 15d-PGJ2 (30 µg·kg−1 i.p.) significantly reduced the: 1) loss of body weight, 2) mortality rate, 3) infiltration of the lung with polymorphonuclear neutrophils (myeloperoxidase activity), 4) oedema formation, and 5) histological evidence of lung injury. Administration of rosiglitazone and 15d-PGJ2 also markedly reduced the nitrotyrosine, PARP and inducible nitric oxide synthase formation. In addition, treatment with the PPAR-γ antagonist bisphenol A diglycidyl ether (1 mg·kg−1 i.p. 30 min before the rosiglitazone or 15d-PGJ2) significantly antagonised the effect of the two PPAR-γ agonists.
These results demonstrate that the two peroxisome proliferator-activated receptor-γ agonists, rosiglitazone and 15-deoxy-Δ12,14-prostaglandin J2, significantly reduce lung injury induced by bleomycin in mice.
- Bleomycin
- 15-deoxy-Δ12,14-prostaglandin J2
- lung injury
- peroxisome proliferator-activated receptor-γ
- rosiglitazone
Pulmonary fibrosis is a progressive interstitial lung disease of unknown aetiology. Pulmonary fibrosis is characterised by inflammatory cell infiltration, fibroblast proliferation and excessive deposition of extracellular matrix proteins in the lung parenchyma 1, 2. The disease most commonly affects middle-aged adults, although infants and children are also affected. Various studies have also indicated that treatment with bleomycin during cancer chemotherapy in humans also induces interstitial fibrosis 3, 4. Several members of the cyclopentenone family of prostaglandins (PGs) possess antineoplastic, antiviral and anti-inflammatory properties 5. PGJ2 is formed by dehydration within the cyclopentane ring of the endogenous prostaglandin, PGD2, and is further metabolised to yield Δ12-PGJ2 and 15-deoxy-Δ12,14-PGJ2 (15d-PGJ2) 5. 15d-PGJ2 is a high-affinity ligand for peroxisome proliferator-activated receptor (PPAR)-γ; a nuclear hormone receptor belonging to the superfamily of ligand-dependent transcription factors related to retinoid, steroid and thyroid hormone receptors, which regulates gene expression by heterodimerising with the retinoid X receptor 6. The actions of PPAR-γ were originally thought to be limited to the control of lipid metabolism and homeostasis. Recent studies, however, have shown that PPAR-γ activation can regulate inflammatory responses and cellular proliferation and differentiation as well as apoptosis 7, 8. Therefore, it is possible that PPAR-γ transrepresses the expression of pro-inflammatory mediators at the transcriptional level by inhibiting nuclear factor-κB (NF-κB), signal transducers and activators of transcription (STAT) 1 and activation protein (AP)-1 signalling 9. There is recent evidence that the cyclopentenone PG 15d-PGJ2 may also play a therapeutic role in conditions associated with inflammation 10, 11. Recently, thiazolidinedione (TZD) derivatives, synthetic PPAR-γ ligands (e.g. rosiglitazone, pioglitazone and roglitazone), have been developed and used as oral antihyperglycaemic agents in the therapy of noninsulin-dependent diabetes mellitus 12, 13. One such compound, rosiglitazone, binds with a high affinity to PPAR-γ 14. All TZDs tested to date, i.e. rosiglitazone, pioglitazone and troglitazone, bind to and activate the PPAR-γ isotype with dissociation constants that parallel their antidiabetic activity in vivo 14, 15. In addition to TZDs, other synthetic compounds have been identified as PPAR-γ activators. It has been also shown that several nonsteroidal anti-inflammatory drugs, such as indomethacin, ibuprofen, fenoprofen, and flufenamic acid, bind to and activate PPAR-γ and promote adipocyte differentiation 16.
The present study investigated the effects of two PPAR-γ agonists in a mouse model of chronic lung injury. Specifically, the effects of the PPAR-γ ligands rosiglitazone (a TZD) and 15d-PGJ2 on the lung injury caused by bleomycin have been investigated. In particular, the effects of the two PPAR-γ agonists (rosiglitazone and 15d-PGJ2) were investigated on the bleomycin-induced: 1) loss of body weight, 2) polymorphonuclear leukocyte infiltration (myeloperoxidase (MPO) activity), 3) tissue oedema (wet:dry ratio), 4) expression of inducible nitric oxide synthase (iNOS; determined immunohistochemically), 5) nitration of tyrosine residues (an indicator of the formation of peroxynitrite), poly(ADP ribose) polymerase (PARP) activation (determined immunohistochemically), and 6) lung damage (histology). Finally, in order to elucidate whether or not the protective effects of rosiglitazone and 15d-PGJ2 are related to activation of the PPAR-γ receptor, it was also investigated whether bisphenol A diglycidyl ether (BADGE), which functional studies have indicated to be an antagonist of PPAR-γ 17, attenuates the protective effects of rosiglitazone and 15d-PGJ2.
MATERIALS AND METHODS
Animals
Male caesarean derived mice (25–35 g; Harlan Nossan, Correzzana, Italy) were housed in a controlled environment and provided with standard rodent food and water. Animal care was in compliance with Italian regulations on the protection of animals used for experimental and other scientific purposes (DM 116192) as well as with European Union regulations (OJ of EC L 358/1 12/18/1986).
Materials
Unless otherwise stated, all compounds were obtained from Sigma-Aldrich Company (Poole, UK). All other chemicals were of the highest commercial grade available. All stock solutions were prepared in nonpyrogenic 0.9% saline (Baxter, Milan, Italy).
Experimental groups
Mice were randomly allocated to the following groups: 1) bleomycin plus vehicle group: mice were subjected to bleomycin-induced lung injury and received the vehicle for rosiglitazone and 15d-PGJ2 (10% dimethylsulphoxide (volume/volume (v/v) i.p. bolus)) every 24 h starting from day 1 (n = 30); 2) BADGE group: same as the bleomycin plus vehicle group except for the administration of BADGE (1 mg·kg−1 i.p. bolus) 30 min prior to vehicle from day 1 (n = 30); 3) rosiglitazone group: same as the bleomycin plus vehicle group except for the administration of rosiglitazone (Alexis, Milan, Italy; 10 mg·kg−1 i.p. bolus) every 24 h starting from day 1 (n = 30); 4) 15d-PGJ2 group: same as the bleomycin plus vehicle group except for the administration of 15d-PGJ2 (30 μg·kg−1 i.p. bolus) every 24 h starting from day 1 (n = 30); 5) BADGE plus rosiglitazone group: same as the rosiglitazone group except for the administration of BADGE (1 mg·kg−1 i.p. bolus) 30 min prior to rosiglitazone (n = 30); 6) BADGE plus 15d-PGJ2 group: identical to the 15d-PGJ2 group except for the administration of BADGE (1 mg·kg−1 i.p. bolus) 30 min prior to 15d-PGJ2 (n = 30); 7) sham plus saline group: sham-operated group, in which identical surgical procedures to the bleomycin group were performed, except that saline was administered instead of bleomycin; 8) sham plus 15d-PGJ2 group: identical to sham plus saline group, except for the administration of 15d-PGJ2 (30 μg·kg−1 i.p. bolus) every 24 h starting from day 1 (n = 30); 9) Sham plus rosiglitazone group: identical to sham plus saline group, except for the administration of rosiglitazone (10 mg·kg−1 i.p. bolus) every 24 h starting from day 1 (n = 30); and 10) sham plus BADGE group: identical to sham plus saline group, except for the administration of BADGE (1 mg·kg−1 i.p. bolus) 30 min prior to the saline. In another set of studies, following bleomycin administration, the various groups of mice (n = 20 per group) were observed for 15 days in order to determine survival differences. The doses of rosiglitazone and 15d-PGJ2 used have previously been reported to reduce the tissue injury caused by inflammation 10, 18.
Induction of lung injury by bleomycin
Mice received a single intratracheal instillation of saline (0.9%) or saline containing bleomycin sulphate (1 mg·kg body weight−1) in a volume of 50 μL and were killed after 15 days by pentobarbitone overdose.
Measurement of fluid content of lung
The lung wet weight was measured after careful excision of extraneous tissues. The lung was exposed for 48 h at 180°C and the dry weight measured. Water content was calculated by subtraction of dry weight from wet weight.
Histological examination
Lung biopsy samples were taken 15 days after injection of bleomycin. Biopsy samples were fixed in 10% (weight/volume (w/v)) PBS-buffered formaldehyde solution for 1 week at room temperature, dehydrated using graded ethanol and embedded in Paraplast (Sherwood Medical, Mahwah, NJ, USA). After embedding in paraffin, the sections were prepared and stained using haematoxylin–eosin or trichrome stain. All sections were studied using light microscopy (Dialux 22; Leitz, Milan, Italy). The severity of fibrosis was semiquantitatively assessed according to the method proposed by Ashcroft et al. 19. Briefly, the grade of lung fibrosis was scored on a scale of 0–8 by examining six randomly chosen fields per sample at a magnification of ×100. The criteria for grading lung fibrosis were as follows: grade 0: normal lung; grade 1: minimal fibrous thickening of alveolar or bronchiolar walls; grade 3: moderate thickening of walls without obvious damage to lung architecture; grade 5: increased fibrosis with definite damage to lung structure and formation of fibrous bands or small fibrous masses; grade 7: severe distortion of structure and large fibrous areas; and grade 8: total fibrous obliteration of fields.
Immunohistochemical localisation of PPAR-γ, iNOS, nitrotyrosine and PARP
Tyrosine nitration, an index of the nitrosylation of proteins by peroxynitrite and/or reactive oxygen species, was determined immunohistochemically as previously described 20. At the end of the experiment, the tissues were fixed in 10% (w/v) PBS-buffered formaldehyde and 8-μm sections prepared from paraffin-embedded tissues. After deparaffinisation, endogenous peroxidase was quenched with 0.3% (v/v) hydrogen peroxide in 60% (v/v) methanol for 30 min. The sections were permeablised with 0.1% (w/v) Triton X-100 in PBS for 20 min. Nonspecific adsorption was minimised by incubating the sections in 2% (v/v) normal goat serum in PBS for 20 min. Endogenous biotin- or avidin-binding sites were blocked by sequential incubation for 15 min with biotin and avidin (DBA, Milan, Italy), respectively. Sections were incubated overnight with anti-nitrotyrosine polyclonal antibody (1:500 in PBS (v/v)), anti-PARP goat polyclonal antibody (1:500 in PBS (v/v)), anti-iNOS polyclonal antibody (1:500 in PBS (v/v)) or anti-PPAR-γ polyclonal antibody (1:500 in PBS (v/v)). Sections were washed in PBS and incubated with secondary antibody. Specific labelling was detected using a biotin-conjugated goat anti-rabbit immunoglobulin G and avidin–biotin peroxidase complex (DBA, Milan, Italy). In order to confirm that the immunoreactions for the nitrotyrosine were specific, some sections were also incubated with the primary antibody (anti-nitrotyrosine) in the presence of excess nitrotyrosine (10 mM). In order to verify the binding specificity for PARP or iNOS, some sections were also incubated with the primary antibody alone (no secondary) or with the secondary antibody alone (no primary). In these situations, no positive staining was found in the sections, indicating that the immunoreactions were positive in all the experiments carried out.
Myeloperoxidase activity
MPO activity, an indicator of polymorphonuclear leukocyte accumulation, was determined as previously described 21. At the specified time following injection of bleomycin, lung tissues were obtained and weighed, each piece homogenised in a solution containing 0.5% (w/v) hexadecyltrimethylammonium bromide in 10 mM potassium phosphate buffer (pH 7) and centrifuged for 30 min at 20,000×g at 4°C. An aliquot of the supernatant was then allowed to react with 1.6 mM tetramethylbenzidine in 0.1 mM hydrogen peroxide. The rate of change in absorbance was measured spectrophotometrically at 650 nm. MPO activity was defined as the quantity of enzyme degrading 1 µmol of peroxide per minute at 37°C and expressed in milliunits per gram of wet tissue.
Statistical evaluation
All data are presented as the mean±sem of n observations. For the in vivo studies, n represents the number of animals studied. In the experiments involving histology or immunohistochemistry, the figures shown are representative of at least three experiments performed on different experimental days. Data sets were examined by one- or two-way ANOVA, and individual group means were then compared using an unpaired t-test. A p-value of <0.05 was considered significant.
RESULTS
Effects of rosiglitazone and 15d-PGJ2 on bleomycin-induced lung injury
Histological examination of the lung sections revealed significant tissue damage by bleomycin (fig. 1b⇓ and f; table 1⇓). Thus, when compared to lung sections taken from saline-treated animals (fig. 1a⇓ and e; table 1⇓), histological examination of the lung sections of mice treated with bleomycin were characterised by extensive inflammatory infiltration by neutrophils, lymphocytes and plasma cells extending through the lung epithelium (fig. 1b⇓ and f; table 1⇓), fibrosis (fig. 1f⇓; table 1⇓) and granulomas in the perivascular region (fig. 1f⇓). Administration of rosiglitazone or 15d-PGJ2 significantly prevented lung inflammation induced by bleomycin administration (fig. 1c⇓, d, g, h). BADGE treatment did not affect the lung injury induced by bleomycin (data not shown). Furthermore, the injection of bleomycin elicited an inflammatory response characterised by the accumulation of water in the lung, as indicated by fluid content (fig. 2⇓) and neutrophil infiltration into the lung tissues (fig. 3⇓). Treatment with either of the two PPAR-γ agonists rosiglitazone and 15d-PGJ2 significantly reduced the fluid content and neutrophil infiltration (figs. 2⇓ and 3⇓). Co-administration of BADGE and rosiglitazone or 15d-PGJ2 significantly blocked the effect of the two PPAR-γ agonists (figs. 2⇓ and 3⇓). BADGE treatment did not reduce the fluid content or neutrophil infiltration (figs. 2⇓ and 3⇓).
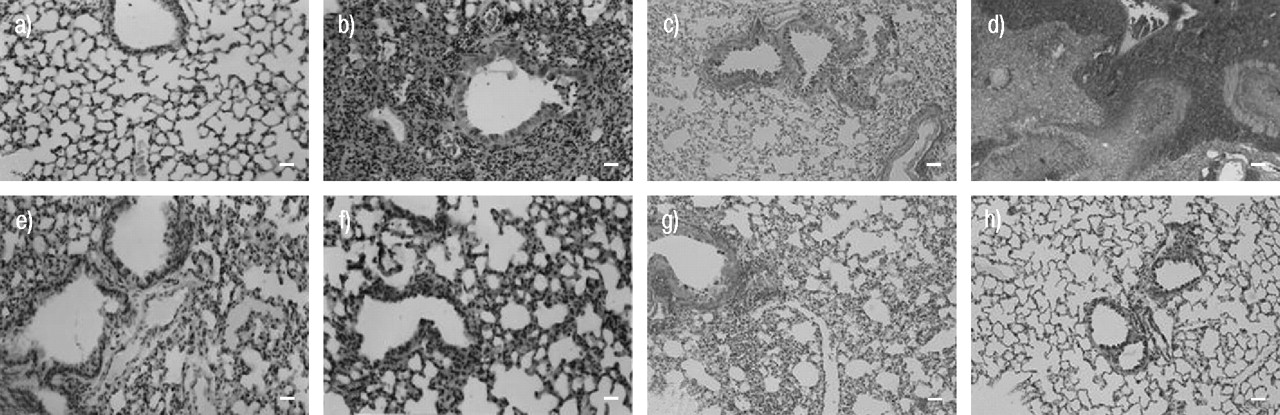
Effects of rosiglitazone and 15-deoxy-Δ12,14-prostaglandin J2 (15d-PGJ2) on lung injury using comparable sections of mouse lung stained with: a, b, e, f) haematoxylin–eosin stain; and c, d, g, h) trichrome stain. a, c) Saline control: normal lung architecture; b, d) bleomycin alone: extensive inflammation with inflammatory cell infiltration and fibrosis (b) and extensive areas of collagen (d); e, g) bleomycin plus rosiglitazone: patchy areas of inflammation with minimal fibrosis (e) and minimal collagen (g); and f, h) bleomycin plus 15d-PGJ2: patchy areas of inflammation with minimal fibrosis (f) and minimal collagen (h). This figure is representative of at least three experiments performed on different experimental days. Internal scale bars = 100 μm, except b) and d) = 200 μm.

Effect of different treatments (□: 10% dimethylsulphoxide; └: 15-deoxy-Δ12,14-prostaglandin J2 (15d-PGJ2); ┘: rosiglitazone;  : bisphenol A diglycidyl ether (BADGE);
: bisphenol A diglycidyl ether (BADGE);  : BADGE/15d-PGJ2;
: BADGE/15d-PGJ2;  : BADGE/rosiglitazone) on the inflammatory response elicited by bleomycin injection and characterised by the accumulation of water in the lung as an indicator of oedema. Data are presented as mean±sem (n = 10 per group). Treatment with either of the two peroxisome proliferator-activated receptor (PPAR)-γ agonists rosiglitazone and 15d-PGJ2 significantly reduced oedema formation. BADGE treatment did not affect the accumulation of water in the lung induced by bleomycin. Co-administration of BADGE and rosiglitazone or 15d-PGJ2 significantly blocked the effect of the two PPAR-γ agonists. **: p<0.01 versus sham; #: p<0.01 versus bleomycin/vehicle; ¶: p<0.01 versus 15d-PGJ2 or rosiglitazone alone.
: BADGE/rosiglitazone) on the inflammatory response elicited by bleomycin injection and characterised by the accumulation of water in the lung as an indicator of oedema. Data are presented as mean±sem (n = 10 per group). Treatment with either of the two peroxisome proliferator-activated receptor (PPAR)-γ agonists rosiglitazone and 15d-PGJ2 significantly reduced oedema formation. BADGE treatment did not affect the accumulation of water in the lung induced by bleomycin. Co-administration of BADGE and rosiglitazone or 15d-PGJ2 significantly blocked the effect of the two PPAR-γ agonists. **: p<0.01 versus sham; #: p<0.01 versus bleomycin/vehicle; ¶: p<0.01 versus 15d-PGJ2 or rosiglitazone alone.

Effect of different treatments (□: 10% dimethylsulphoxide; └: 15-deoxy-Δ12,14-prostaglandin J2 (15d-PGJ2); ┘: rosiglitazone;  : bisphenol A diglycidyl ether (BADGE);
: bisphenol A diglycidyl ether (BADGE);  : BADGE/15d-PGJ2;
: BADGE/15d-PGJ2;  : BADGE/ rosiglitazone) on myeloperoxidase (MPO) activity in the lungs of bleomycin-treated mice. Data are presented as mean±sem (n = 10 per group). MPO activity in the lungs of bleomycin-treated mice was significantly increased in comparison to sham-operated mice. Rosiglitazone and 15d-PGJ2 treatment reduced the bleomycin-induced increase in MPO activity. BADGE treatment did not affect the increase in MPO activity in the lung induced by bleomycin. Co-administration of BADGE and rosiglitazone or 15d-PGJ2 significantly blocked the effect of the two peroxisome proliferator-activated receptor-γ agonists. **: p<0.01 versus sham; #: p<0.01 versus bleomycin/vehicle; ¶: p<0.01 versus 15d-PGJ2 or rosiglitazone alone.
: BADGE/ rosiglitazone) on myeloperoxidase (MPO) activity in the lungs of bleomycin-treated mice. Data are presented as mean±sem (n = 10 per group). MPO activity in the lungs of bleomycin-treated mice was significantly increased in comparison to sham-operated mice. Rosiglitazone and 15d-PGJ2 treatment reduced the bleomycin-induced increase in MPO activity. BADGE treatment did not affect the increase in MPO activity in the lung induced by bleomycin. Co-administration of BADGE and rosiglitazone or 15d-PGJ2 significantly blocked the effect of the two peroxisome proliferator-activated receptor-γ agonists. **: p<0.01 versus sham; #: p<0.01 versus bleomycin/vehicle; ¶: p<0.01 versus 15d-PGJ2 or rosiglitazone alone.
1 Histological scoring of lung fibrosis
Effects of rosiglitazone and 15d-PGJ2 on bleomycin-induced iNOS expression, nitrotyrosine formation and PARP activation
Immunohistochemical analysis of lung sections obtained from bleomycin-treated mice revealed positive staining for iNOS, localised mainly in plasma cell and lymphocytes (fig. 4b⇓). In contrast, no staining for iNOS was found in the lungs of bleomycin-treated mice that had been pretreated with rosiglitazone (fig. 4c⇓) or 15d-PGJ2 (fig. 4d⇓). Staining was absent in lung tissue obtained from the sham group (fig. 4a⇓). Immunohistochemical analysis of lung sections obtained from mice treated with bleomycin also revealed positive staining for nitrotyrosine, localised mainly in the nuclei of inflammatory cells (fig. 5b⇓). In contrast, no positive staining for nitrotyrosine was found in the lungs of bleomycin-treated mice that had been treated with rosiglitazone (fig. 5c⇓) or 15d-PGJ2 (fig. 5d⇓). Immunohistochemical analysis of lung sections obtained from mice treated with bleomycin also revealed positive staining for PARP, localised mainly in plasma cells and lymphocytes (fig. 6b⇓). In contrast, no staining for PARP was found in the lungs of bleomycin-treated mice that had been treated with rosiglitazone (fig. 6c⇓) or 15d-PGJ2 (fig. 6d⇓). There was no staining for either nitrotyrosine or PARP in lungs obtained from the sham group of mice (figs. 5a⇓ and 6a⇓). Co-administration of BADGE and rosiglitazone or 15d-PGJ2 significantly blocked the effect of the two PPAR-γ agonists (data not shown).
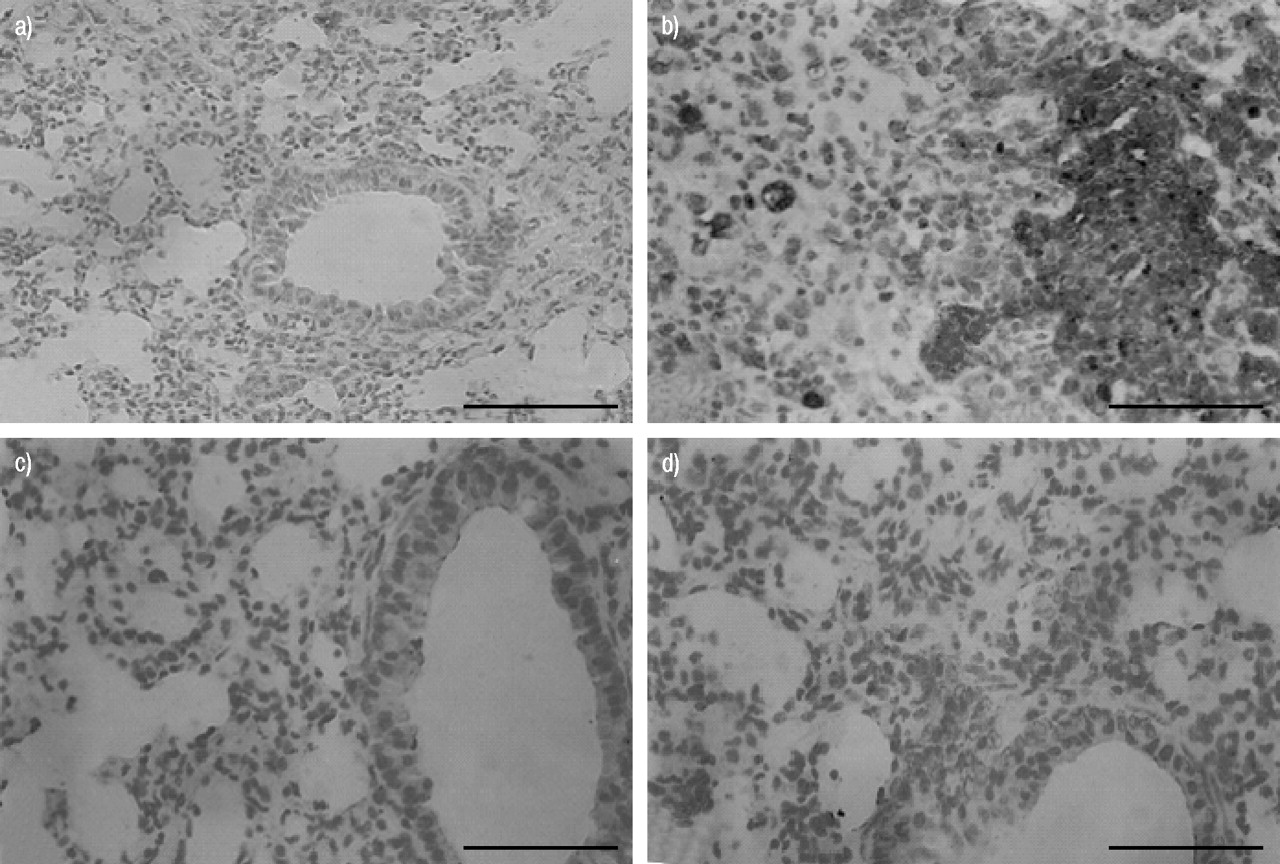
Immunohistochemical localisation of inducible nitric oxide synthase (iNOS) in the lung. a) No positive staining was observed in the lung section from sham-treated mice. b) After bleomycin injection, positive staining for iNOS was localised mainly in plasma cells and lymphocytes. c, d) There was a marked reduction in immunostaining in the lungs of bleomycin-treated mice that had been treated with rosiglitazone (c) and 15-deoxy-Δ12,14-prostaglandin J2 (d). This figure is representative of at least three experiments performed on different experimental days. Scale bar = 22 µm.

Immunohistochemical localisation of nitrotyrosine in the lung. a) No positive staining was observed in the lung section from sham-treated mice. b) After bleomycin injection, positive staining for nitrotyrosine was localised mainly in the nuclei of inflammatory cells. c, d) There was a marked reduction in immunostaining in the lungs of bleomycin-treated mice that had been treated with rosiglitazone (c) and 15-deoxy-Δ12,14-prostaglandin J2 (d). This figure is representative of at least three experiments performed on different experimental days. Scale bar = 22 µm.
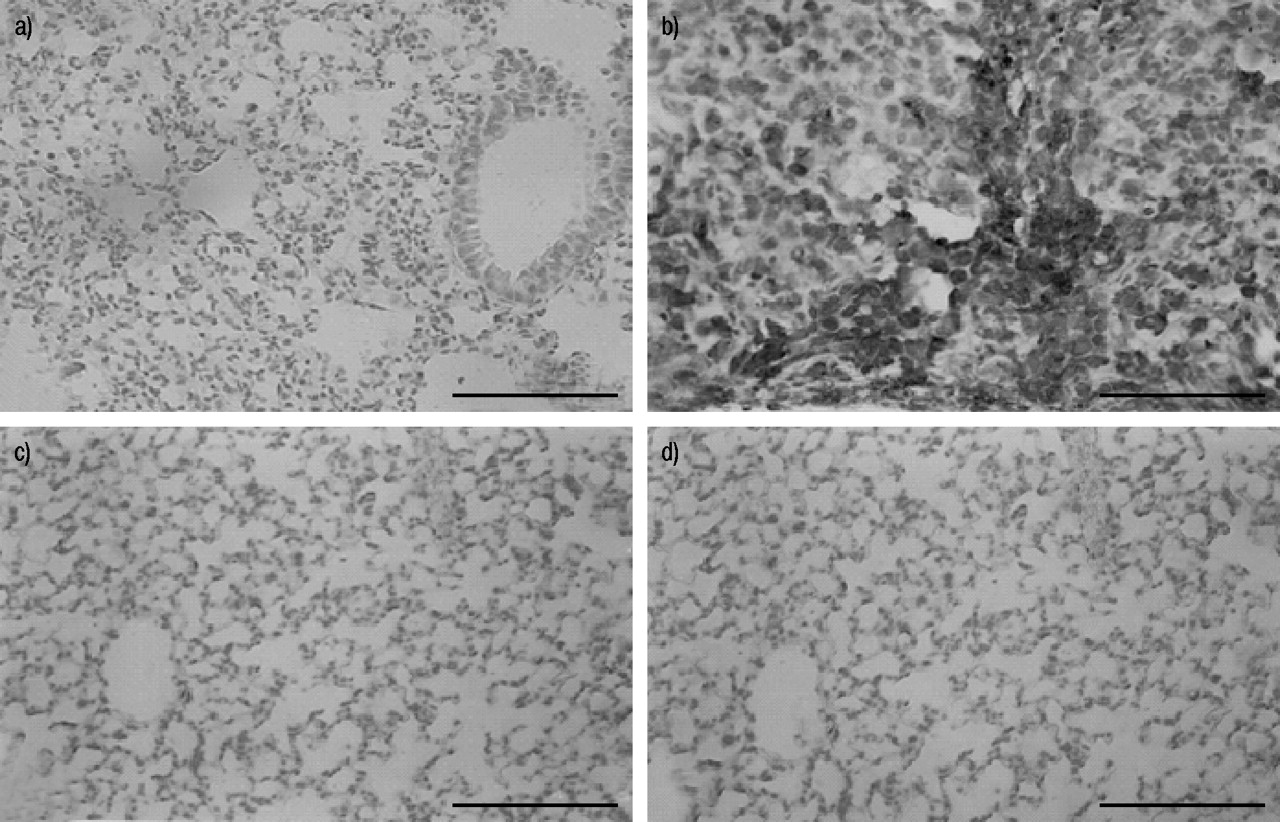
Immunohistochemical localisation of poly(ADP ribose) polymerase (PARP) in the lung. a) No positive staining was observed in the lung section from sham-treated mice. b) After bleomycin injection, positive staining for PARP was localised mainly in plasma cells and lymphocytes. c, d) There was a marked reduction in immunostaining in the lungs of bleomycin-treated mice that had been treated with rosiglitazone (c) and 15-deoxy-Δ12,14-prostaglandin J2 (d). This figure is representative of at least three experiments performed on different experimental days. Scale bar = 22 µm.
BADGE treatment alone did not affect the iNOS expression, nitrotyrosine formation and PARP activation induced by bleomycin administration (data not shown).
Effects of rosiglitazone and 15d-PGJ2 on changes in body weight and survival rate
In vehicle-treated mice, the severe lung injury caused by bleomycin administration was associated with a significant loss of body weight (fig. 7⇓). Treatment with either of the two PPAR-γ agonists rosiglitazone and 15d-PGJ2 significantly reduced the loss of body weight (fig. 7⇓). The survival of animals was monitored for 15 days. Bleomycin-treated mice that had received vehicle developed severe lung injury and 40 and 50% of these animals died within 10 and 15 days, respectively, after bleomycin administration. In contrast, none of the mice that had been treated with either of the two PPAR-γ agonists rosiglitazone and 15d-PGJ2 died (fig. 8⇓). Co-administration of BADGE and rosiglitazone or 15d-PGJ2 significantly blocked the beneficial effects of the two PPAR-γ agonists (figs. 7⇓ and 8⇓). BADGE treatment did not affect the body weight loss and mortality induced by bleomycin administration (figs. 7⇓ and 8⇓).

Effect of rosiglitazone and 15-deoxy-Δ12,14-prostaglandin J2 (15d-PGJ2) treatment on body weight following bleomycin-induced lung injury (with: ▪: 10% dimethylsulphoxide; •: 15d-PGJ2; ♦: rosiglitazone; □: bisphenol A diglycidyl ether (BADGE); ○: BADGE/15d-PGJ2; ⋄: BADGE/rosiglitazone). Body weight was recorded immediately before bleomycin administration and daily for the whole experimental period. Data are presented as mean±sem (n = 10 per group). Rosiglitazone and 15d-PGJ2 treatment significantly prevented loss of body weight. BADGE treatment did not affect the body weight loss induced by bleomycin. Co-administration of BADGE and rosiglitazone or 15d-PGJ2 significantly blocked the effect of the two PPAR-γ agonists. **: p<0.01 versus bleomycin/vehicle; #: p<0.01 versus 15d-PGJ2 or rosiglitazone alone.
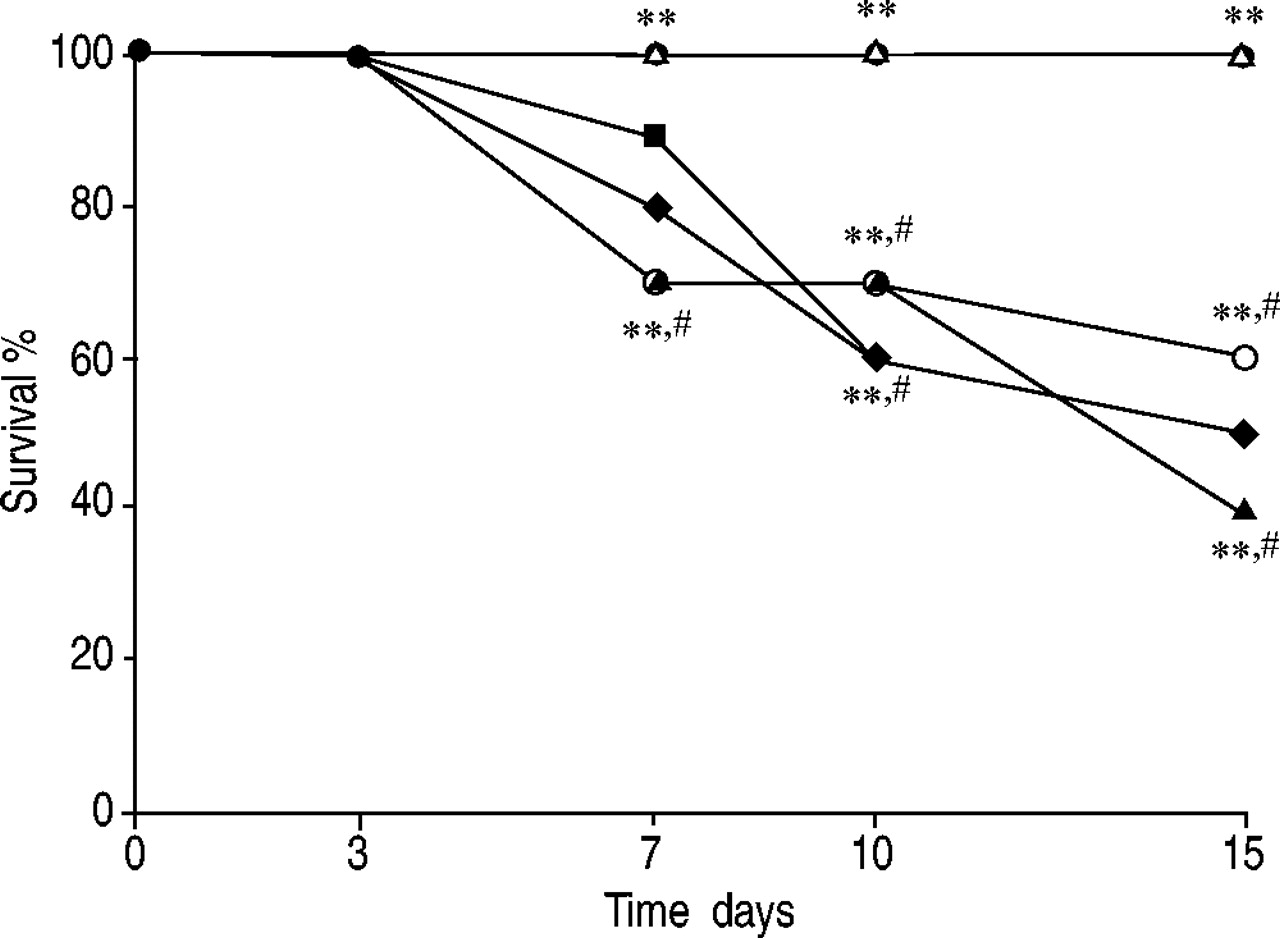
Effect of rosiglitazone and 15-deoxy-Δ12,14-prostaglandin J2 (15d-PGJ2) treatment on bleomycin-induced mortality (with: ♦: 10% dimethylsulphoxide; •: 15d-PGJ2; ▵: rosiglitazone; ▪: bisphenol A diglycidyl ether (BADGE); ○: BADGE/15d-PGJ2; ▴: BADGE/rosiglitazone). Data are presented as mean±sem (n = 10 per group). Survival was significantly improved in rosiglitazone- and 15d-PGJ2-treated mice in comparison to the high mortality rate of the bleomycin-treated mice. BADGE treatment did not affect the mortality induced by bleomycin. Co-administration of BADGE and rosiglitazone or 15d-PGJ2 significantly blocked the effect of the two PPAR-γ agonists. **: p<0.01 versus bleomycin/vehicle; #: p<0.01 versus 15d-PGJ2 or rosiglitazone alone.
Characterisation of PPAR-γ in the lung
In order to confirm that PPAR-γ expression occurs at demonstrable levels in the lung following bleomycin-induced injury, PPAR-γ expression was assessed immunohistochemically. Immunohistochemical analysis of lung sections obtained from bleomycin-treated mice revealed positive staining for PPAR-γ, localised mainly in pneumocyte nuclei as well as in inflammatory cells (fig. 9⇓). Thus PPAR-γ is expressed at significant levels in lung tissue after bleomycin-induced injury.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Peroxisome proliferator-activated receptor (PPAR)-γ expression. Immunohistochemical analysis of lung sections obtained from bleomycin-treated mice revealed positive staining for PPAR-γ, localised mainly in pneumocyte nuclei as well as in inflammatory cells. This figure is representative of at least three experiments performed on different experimental days. Scale bar = 22 µm.
DISCUSSION
Idiopathic pulmonary fibrosis is a progressive interstitial lung disease of unknown aetiology. The disease most commonly affects middle-aged adults, although infants and children are also affected. It is characterised by excessive deposition of extracellular matrix in the lung interstitium. The pathological features of inflammation and fibrosis are well appreciated but little is known about its aetiology and pathogenesis 22, 23. To date, there is no satisfactory treatment for idiopathic pulmonary fibrosis. Various anti-inflammatory agents, such as corticosteroids 24, colchicines 25 and cytotoxic agents such as azathioprine 26 and cyclophosphamide 27, have been used alone or in combination to treat the disease. However, less than a third of patients respond to treatment with corticosteroids and/or cytotoxic therapy.
Only recent studies have implicated the various PPAR isoforms in inflammation 28, 29. In particular, both PPAR-α and PPAR-γ have been reported to regulate the inflammatory response 30; however, the contribution of PPAR-γ to the inflammatory response is unclear 29. PPAR-γ is the molecular target of the TZD class of antidiabetic drugs, which include rosiglitazone and pioglitazone 31. PPAR-γ is prominently expressed in activated monocytes and tissue macrophages, including the foam cells of atherosclerotic lesions, in vivo32. Multiple functions have been proposed for PPAR-γ in inflammation, but how the PPAR-γ signalling pathway might affect the development of acute inflammation remains unclear. Therefore, the present study evaluated the effect of two PPAR-γ agonists in bleomycin-induced lung injury, which has been well characterised by various groups as a useful model for assessing the contribution of mediators involved in lung fibrosis. The current study provides evidence that the two PPAR-γ agonists attenuate: 1) development of bleomycin-induced lung injury, 2) infiltration of the lung with inflammatory cells, 3) the degree of nitrosative stress and PARP formation in the lung, and 4) expression of iNOS. All of these findings support the view that the two PPAR-γ agonists attenuate the degree of bleomycin-induced lung injury in mice. In order to confirm whether this effect of rosiglitazone and 15d-PGJ2 rosiglitazone occurs via activation of PPAR-γ, it has been investigated whether the PPAR-γ antagonist BADGE attenuates the observed protective effect of rosiglitazone and 15d-PGJ2. It was demonstrated here that BADGE does, indeed, attenuate the protective effects of rosiglitazone and 15d-PGJ2 in mice subjected to bleomycin-induced lung injury. Thus the present authors suggest that the protective effects of rosiglitazone and 15d-PGJ2 are secondary to activation of PPAR-γ.
A number of recent studies have demonstrated that the recruitment of cells into an area of inflammation may be mediated not only by the complement component C5a, leukotrienes, platelet-activating factor and bacterially derived peptides but also by a novel group of small proteins with relatively specific chemotactic activity for leukocyte subpopulations. In the present study, it is reported that, in the lung tissue of bleomycin-treated mice that received the two PPAR-γ agonists, a significant reduction in leukocyte infiltration, as assessed by the specific granulocyte enzyme MPO, with moderation of tissue damage, as evaluated by histological examination, was observed. Neutrophils that are recruited into tissue can contribute to tissue destruction by the production of reactive oxygen metabolites, granule enzymes and cytokines that further amplify the inflammatory response by their effects on macrophages and lymphocytes 33. Neutrophils have been shown to express PPAR-γ 34, suggesting a potential role for this transcription factor in neutrophil function. Other studies have also demonstrated that PPAR-γ ligands may modulate leukocyte–endothelial cell interactions during inflammation through regulation of endothelial adhesion molecules. In the present study, the presence of PPAR-γ (assessed immunohistochemically) in lungs from bleomycin-treated mice has clearly been confirmed. Furthermore, the discovery that the tissue damage induced by bleomycin in vehicle-treated mice was associated with intense immunostaining due to nitrotyrosine formation also suggested that a lung structural alteration had occurred, most probably due to the formation of highly reactive nitrogen derivatives. Recent evidence indicates that bleomycin is a well-known cause of intracellular oxidative stress, and several findings in the present study suggest that extracellular oxidative stress may also play a role in the pathogenesis of bleomycin-induced lung injury 35. Therefore, in the present study, it is clearly demonstrated that the two PPAR-γ agonists prevent induction of iNOS and formation of peroxynitrite.
Reactive oxygen species produce strand breaks in DNA, which trigger energy-consuming DNA repair mechanisms and activate the nuclear enzyme PARP, resulting in depletion of its substrate, oxidised nicotinamide adenine dinucleotide (NAD), in vitro and a reduction in the rate of glycolysis. As NAD functions as a cofactor in glycolysis and the tricarboxylic acid cycle, NAD depletion leads to a rapid fall in intracellular ATP levels. This process has been termed “the PARP suicide hypothesis” 36. There is recent evidence that activation of PARP may also play an important role in inflammation 37. It is demonstrated here that PPAR-γ agonist treatment reduced activation of PARP during bleomycin-induced lung injury. Thus, the present authors propose that the anti-inflammatory effects of rosiglitazone and 15d-PGJ2 may be, at least in part, due to prevention of PARP activation. Several cellular mechanisms, including the mode of gene regulation and signal transduction, may account for the anti-inflammatory effect of PPAR-γ ligands. Recently, it has been shown that PPAR-γ ligands may act at the transcriptional level, at least in part, through inhibition of AP-1 and NF-κB activity 38.
Although the exact mechanisms remain unclear, activated PPAR-γ could downregulate AP-1, NF-κB and STAT activity by titration of essential transcription cofactors, such as cAMP-response element-binding protein/p300 and steroid receptor coactivator-1 39. PPAR may also antagonise AP-1 and NF-κB activity through protein–protein interaction. Such a mechanism has been shown to be utilised by the retinoic acid and glucocorticoid receptors with AP-1or NF-κB 40. NF-κB has been shown to activate, via transcription, the genes encoding the pro-inflammatory iNOS.
In conclusion, the present study provides the first evidence that rosiglitazone, a ligand of PPAR-γ, causes a substantial reduction in the lung injury induced by bleomycin administration. In addition, the protective effect of 15d-PGJ2 has been confirmed in this experimental model of lung injury. Thus, it is demonstrated here that the mechanisms underlying the protective effects of rosiglitazone and 15d-PGJ2 are dependent on activation of PPAR-γ. The activation of PPAR-γ by rosiglitazone and 15d-PGJ2, in turn, results in a reduction in: 1) expression of inducible nitric oxide synthase and nitration of proteins by peroxynitrite; 2) poly(ADP ribose) polymerase activation; 3) inflammatory cell infiltration; and 4) lung injury. These findings support the potential use of PPAR-γ ligands as therapeutic agents in the therapy of conditions associated with lung inflammation.
Acknowledgments
The authors would like to thank G. Pergolizzi and C. La Spada for their excellent technical assistance, C. Cutrona for secretarial assistance and V. Malvagni for editorial assistance with the manuscript.
- Received April 27, 2004.
- Accepted September 14, 2004.
- © ERS Journals Ltd
References









