





Abstract
Alveolar epithelial type I cells cover most of the internal surface area of the lungs.
Ultrastructural studies demonstrate that alveolar epithelial type I cell damage is frequently observed in acute and chronic lung diseases.
This article discusses the use of cell-selective proteins as markers for the investigation of injury and repair of the alveolar epithelium. The utility of proteins specific to alveolar epithelial type I cells as diagnostic markers of alveolar epithelial injury in acute lung injury is considered, and expression of proteins selective for alveolar epithelial type I cells in lungs following injury and in fibrosis are discussed.
- Alveolar epithelial type I cell
- cell differentiation
- fibrosis
- marker
- primary acute respiratory distress syndrome
- pulmonary injury
M.C. McElroy is funded by the Norman Salvesen Emphysema Research Fund (Edinburgh, UK) and M. Kasper by the Deutsche Forschungsgemeinschaft (SFB 13) (Bonn, Germany).
Epithelial damage is a major hallmark of many life-threatening pulmonary diseases, including acute lung injury (ALI) or acute respiratory distress syndrome (ARDS), severe acute respiratory syndrome, cystic fibrosis and idiopathic pulmonary fibrosis (IPF) 1–4. In addition, alveolar epithelial injury followed by abnormal epithelial repair appears to be a key pathological feature of IPF 4.
Two cell types populate the alveolar epithelium in normal adult lungs, alveolar epithelial type I (ATI) and alveolar epithelial type II (ATII) cells. ATI cells cover ∼95% of the internal surface area of the lung. They are branched cells with multiple apical surfaces that extend into adjacent alveoli 5. The apical surface area of ATI cells is large in comparison with most cells (i.e. ∼5,000 µm2 for human ATI cells), yet they are very thin cells (i.e. 0.2 µm in depth) 5, 6. The gas exchange barrier is composed of ATI and endothelial cells joined by fused basement membranes. ATI cells are important in the regulation of alveolar fluid balance and surfactant secretion by ATII cells in response to stretch 7–10. ATII cells cover the remaining 2–5% of the lung's surface area. They are cuboidal cells situated between ATI cells, and contain characteristic lamellar bodies and apical microvilli 5. ATII cells have many known functions, including the production, secretion and reuptake of pulmonary surfactant 11, regulation of alveolar fluid in normal lungs and during the resolution of pulmonary oedema 12, and synthesis and secretion of immunomodulatory proteins important for host defence, such as surfactant proteins A and D 13, 14.
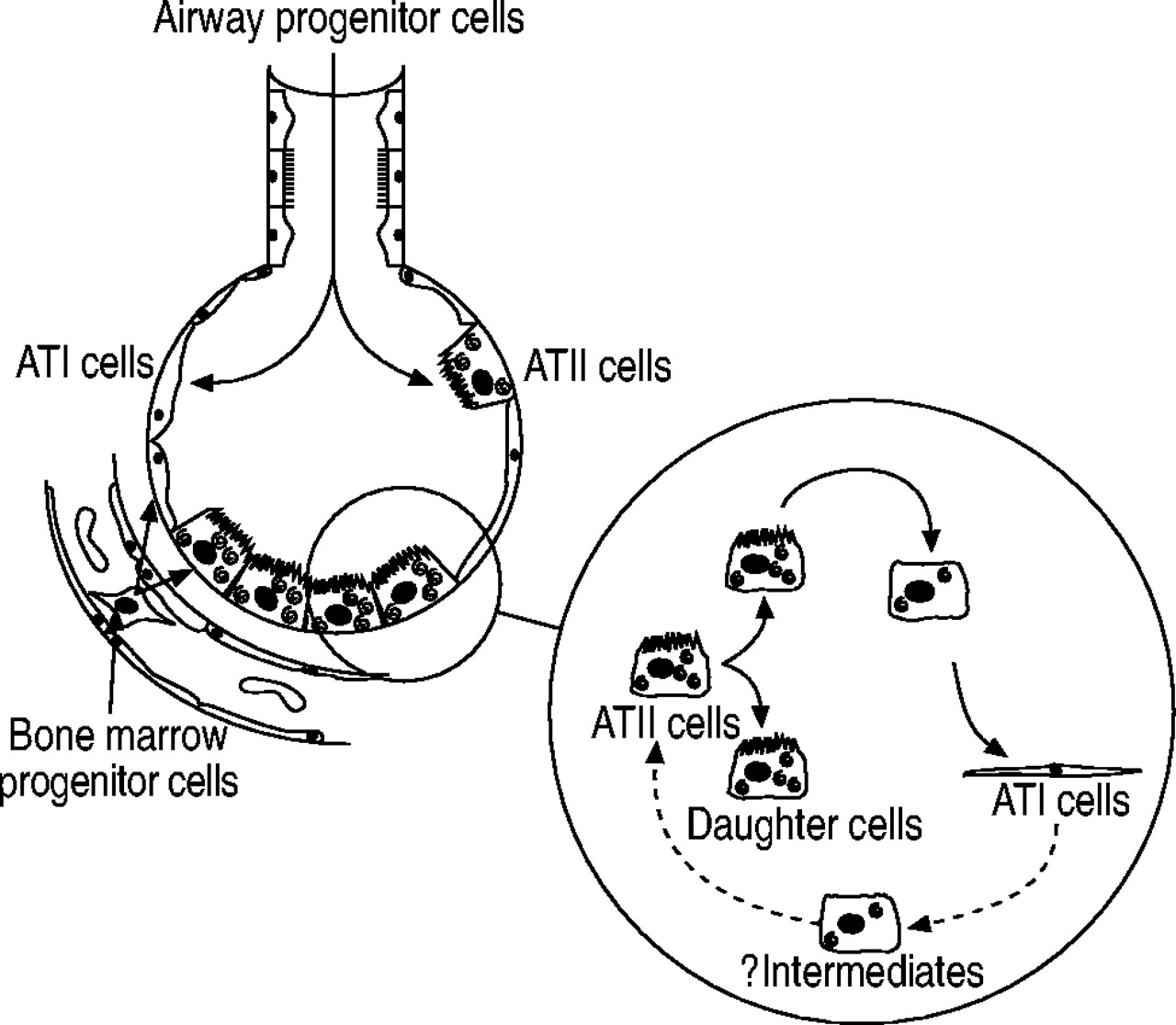
Damaged ATI cells are replaced by ATII cells 15–19 (reviewed in 19) (fig. 1⇓). The studies of Evans and coworkers 15, 20, in particular, provide the foundation of current knowledge concerning the response of the alveolar epithelium to injury. They examined ultrastructural changes to the alveolar epithelium following exposure to the oxidant gas nitrogen dioxide for 48 h. In alveoli adjacent to terminal bronchioles, ATI cells were maximally damaged after 8–12 h 20. After 48 h of exposure, the denuded basement membranes were repopulated by cuboidal epithelial cells, some of which were ATII cells. Evans et al. 15 followed the fate of proliferating ATII cells (after 48 h of NO2 exposure) using a combination of autoradiography and electron microscopic analysis. Proliferating cells were labelled with tritiated thymidine, which is incorporated into newly synthesised DNA during the S phase of the cell cycle. After 48 h of NO2 exposure, most labelled cells were ATII cells; however, following afurther 48-h recovery period, the number of labelled ATII cells had decreased, whereas the number of labelled ATI cells had increased. In addition, over the 48-h recovery period, there was a transient increase in label associated with alveolar epithelial cells with characteristics of both ATI and ATII cells (i.e. ATII-like cells but spread out over the basement membrane or ATI-like cells containing occasional microvilli and lamellar bodies) 15. These studies demonstrate three important aspects of alveolar epithelial injury and repair (in response to NO2). First, the loss of ATI cells can precede ATII cell proliferation. Secondly, alveolar epithelial repair involves the transformation ATII cells (or a subset thereof) into new ATI cells. Thirdly, ATII cells transforming to new ATI cells do so via intermediate cell types.

Alveolar epithelial repair. Following injury, the alveolar epithelium is repaired by the proliferation and transdifferentiation of alveolar epithelial type II (ATII) cells, via intermediate cells, into new alveolar epithelial type I (ATI) cells (solid arrows). Injured alveolar epithelial cells may also be replaced by bone marrow and airway progenitor cells. The role of ATI cells in alveolar epithelial repair is still speculative (dotted arrows).
More recent studies have refined the role of ATII cells in ATI cell repair and raised alternative mechanisms of ATI cell repair (fig. 1⇑) 21–24. There may be subpopulations of ATII cells with different proliferative capabilities following injury 24. However, the potential for these putative ATII cell subpopulations to transdifferentiate into ATI cells is unknown. Damaged ATI cells may also be replaced by bone marrow and nonciliated bronchiolar epithelial progenitor cells 21, 22. It has been assumed for a long time that ATI cells were terminally differentiated. However, experiments designed to investigate the role of mechanical load in foetal lung development suggest that ATI cells have the potential to transdifferentiate into ATII cells 23. It is not known whether ATI cells are capable of replacing damaged ATII cells in injured adult lungs.
Investigating ATI cell injury and repair is difficult because they are not easily recognised using light microscopy. To date, most studies have relied on visualisation of ATI cells, and, in particular, necrosis of ATI cells, using electron microscopy. A number of proteins have been identified which are expressed by ATI cells but not ATII cells (reviewed in 25). The present review focuses on the application of ATI-cell-selective proteins as markers for the investigation of injury to and repair of ATI cells in experimental models and human lung diseases. First, known ATI-cell-selective proteins are described (see also 25). Secondly, the utility of ATI-cell-specific proteins in biochemical assays for quantification of the extent of ATI cell necrosis is discussed. Thirdly, the significance of changes in ATI-cell-selective proteins following injury and in lung fibrosis is evaluated.
Proteins expressed by alveolar epithelial type I cells
In the following section, the ATI-cell-selective proteins have been categorised into three groups (table 1⇓). Group I contains proteins that are specifically expressed by ATI cells in the lung (ATI-cell-specific). Group II contains proteins expressed by ATI, and other lung cells, but not ATII cells (ATI-cell-selective), and group III contains proteins either expressed more abundantly by ATI than ATII cells or for which there is insufficient evidence to place them in group II (ATI-cell-selective/associated).
Alveolar epithelial type I (ATI) cell-specific and -associated proteins in normal adult lung
Group I. alveolar epithelial type I cell-specific proteins
Rat type I cell 40-kDa protein/type I cell alpha protein
Rat type I cell 40-kDa protein (RTI40)/type I cell alpha (TIα) protein is an apical membrane protein expressed exclusively by ATI cells in the lung (fig. 2⇓) 26, 52, 53. A monoclonal antibody directed against RTI40/TIα protein was used to identify and characterise the rat gene TIα in a bacterial expression library 53. The TIα gene encodes a membrane protein predicted to have a large extracellular domain, a transmembrane region and a short cytoplasmic tail 53. RTI40/TIα protein is also detected in nonpulmonary tissues, including the choroid plexus and ciliary epithelium of the eye 54. RTI40/TIα protein is homologous to a number of other proteins, including OTS-8 (mouse phorbol ester-treated osteoblast cell line 55), 38-kDa glycoprotein (gp38) (mouse thymic epithelial cells 56), E11 (rat lymphatic endothelial cells and osteoblasts 57), podoplanin (rat kidney podocytes 58), 40-kDa glycoprotein (gp40) (Madin-Darby canine kidney cells 59), PA2.26 antigen (mouse keratinocytes during epidermal carcinogenesis 60) and Aggrus (human colorectal tumours 27). Mouse and rat RTI40/TIα protein are 78% homologous on the basis of deduced amino acid sequences 53.

Use of alveolar epithelial type I (ATI) cell-specific antibodies to visualise rat ATI cells using immunofluorescence microscopy at a) low and b) high power. ATI cells are stained green with a monoclonal antibody directed against rat type I cell 40−kDa protein (RTI40)/type I cell alpha (TIα) protein. Alveolar epithelial type II cells and Clara cells are stained red with the MMC4 monoclonal antibody (arrow). Br: bronchiole; BV: blood vessel. *: airspace. (From 51.)
Human homologues of RTI40/TIα protein have been cloned and are called hTIα-2 and gp36 61, 62. hTIα-2 is 50% identical to TIα at the nucleotide level. Northern blot analysis of messenger ribonucleic acid (mRNA) demonstrates that the transcript for hTIα-2 is present in lung and not in other selected tissues (e.g. kidney and small intestine) 61. gp36 is 66% identical to TIα at the nucleotide level. Sequence alignment demonstrates that the greatest variability between RTI40/TIα protein and gp36 occurs in the extracellular domain, which might explain why antibodies that recognise rat ATI cells do not also recognise human ATI cells 62. Although a monoclonal antibody raised against gp36 is reported to recognise the alveolar epithelium, there is insufficient data at present to determine whether gp36 is ATI-cell-specific in human lungs 62.
Mice with targeted mutations in the TIα locus die at birth due to respiratory failure 63, 64. Histological analysis of the lungs demonstrates that mice deficient in RTI40/TIα protein exhibit fewer ATI cells, narrower and irregular airspaces, and defective formation of alveolar saccules in comparison to wild-type litter mates 63. In addition, there is abnormally high expression of proliferation markers associated with the alveolar epithelium and mesenchymal cells (e.g. fibroblasts) in the distal lung 63. These data demonstrate that the loss of TIα is important, although not absolutely required, for the formation of ATI cells during the late stages of gestation in the mouse. These data suggest that TIα, ATI cells or both regulate distal airway cell proliferation during lung development.
The role of RTI40/TIα protein in normal adult lung is unknown. RTI40/TIα protein is a sialoglycoprotein 52, 59, 62. Highly negatively charged proteins, such as sialoglycoproteins, are relatively resistant to proteases. Therefore, RTI40/TIα protein may act as a physical barrier and protect the surface of ATI cells from proteases 62. The overexpression of TIα in endothelial cells and keratinocytes in culture promotes phenotypic changes (i.e. long thin cell extensions) and increases the migration rate in wounding experiments 60, 64. In adult lungs, RTI40/TIα protein may, therefore, be important for initiating and maintaining the extended and flattened ATI cell phenotype. A similar role has been proposed for RTI40/TIα in protein kidney podocytes 58. However, other recognised functions of RTI40/TIα protein are harder to explain. RTI40/TIα protein has also been identified as a platelet aggregation-inducing factor expressed by colorectal tumours 27. It is not known whether RTI40/TIα protein on the surface of ATI cells facilitates platelet aggregation.
Human type I cell 56-kDa protein
Dobbs et al. 29 have also generated an antibody that specifically reacts with ATI cells in human lungs, human type I cell 56-kDa protein (HTI56). This antibody has been used to quantify ATI cell injury in patients with established ARDS (see later). It is not known whether HTI56 and RTI40 are homologous. At present, there are no data on the biological or functional properties of HTI56.
Group II. alveolar epithelial type I cell-selective proteins
Immunoelectron microscopy has been used to demonstrate that caveolin-1 and -2 30, 31, cytochrome P450 2B1 34, aquaporin (AQP) 5 33, 65 and the α2-isoform of sodium/potassium-exchanging adenosine triphosphatase (Na+/K+-ATPase) 32 are expressed by ATI but not ATII cells (table 1⇑).
Caveolin-1 and -2
Caveolae are small flask-shaped microvesicles found in abundance in ATI and endothelial cells in the lung. The principle protein component of caveolae is caveolin-1, which is required for the biogenesis of these organelles 66, 67 (reviewed in 68, 69). In the lung, caveolin-2 is coexpressed with caveolin-1 (70 and references therein). Caveolae are also enriched with cholesterol and sphingolipids and may serve as specific microdomains or “rafts” in which cell signalling cascades are regulated (reviewed in 69). In particular, caveolin-1 may negatively regulate proliferation and apoptosis, and may, therefore, act as a tumour suppressor 69, 71. Mice deficient in either caveolin-1 or -2 are born healthy and are able to reproduce 66, 67, 70. However, by 4–5 months of age, they develop a lung pathology that resembles fibrosis. Specifically, caveolin-1 and -2 knockout mice show smaller airspaces and thickened alveolar walls 66, 67, 70, 72. The increased alveolar wall thickness is due to endothelial hyperproliferation and increased deposition of extracellular matrix proteins 69–72. Both types of knockout mice are also exercise-intolerant in comparison with their wild-type litter mates 66, 67, 70. The pulmonary defect may be caused solely by caveolin-2 deficiency 70. Caveolin-1-deficient mice show a dramatic reduction in caveolin-2 levels 66, 67, whereas caveolin-2 null mice show almost normal levels of caveolin-1 70. It has been hypothesised that caveolin-2 might be required for the mitigating mechanical forces that stimulate lung cell proliferation 70.
α2-Isoform of sodium/potassium-exchanging adenosine triphosphatase
The alveoli have to be maintained essentially fluid-free in order to ensure efficient gas exchange (reviewed in 12). In addition, the upregulation of fluid absorption following acute lung injury is an important factor in recovery from ALI or ARDS 12. Fluid is removed from the airspaces by active transport of sodium across the epithelium by a basolateral Na+/K+-ATPase. The resulting osmotic gradient promotes reabsorption of water 12. Until recently, it was believed that ATII cells were the most important cell type in the regulation of alveolar fluid clearance. However, it has now been shown that ATI cells contain apical sodium channels and can actively transport sodium ions 8, 32, 73. Immunoelectron microscopy suggests that the α2-isoform of Na+/K+-ATPase is expressed in ATI but not ATII cells and accounts for a significant percentage of basal alveolar fluid reabsorption 32.
Aquaporin 5
AQP5 is a water channel protein located in the apical membrane of ATI cells and bronchiolar epithelial cells in the lung 33, 73. AQP5-deficient mice demonstrate that AQP5 is important for the movement of water from the airspaces to the capillary bed in response to an osmotic gradient generated from the vascular bed 74. However, AQP5 does not appear to be important in water absorption out of the airspaces under normal conditions or under conditions in which alveolar fluid clearance is upregulated by specific agonists 74. Nor is AQP5 important in lung extravascular fluid accumulation in acute injury 75. It has been proposed that AQP5 may be important in the regulation of ATI cell volume 76.
Cytochrome P450 2B1 and carboxypeptidase M
Both cytochrome P450 2B1 and carboxypeptidase M are associated with ATI but not ATII cells in the lung 35, 77. Cytochrome P450 2B1 is also present in nonciliated bronchiolar epithelial cells and carboxypeptidase M in macrophages 34, 35. The specific role of these enzymes in ATI cell function is unknown.
Group III. Alveolar epithelial type I cell-selective/associated proteins
Intercellular adhesion molecule-1
Alveolar epithelial intercellular adhesion molecule-1 (ICAM-1) may specifically regulate alveolar macrophage phagocytic activity in the lung 78 or remodelling of the alveolar epithelium following injury 1. Immunoelectron micrographs demonstrate that ICAM-1 is present on the apical surface of ATI cells, and, in particular, that there is abundant expression on surfaces near cell junctions 36. ICAM-1 is also detected at lower levels on the surface of capillary endothelial cells 78. Immunoelectron microscopic studies demonstrate low levels of ICAM-1 on ATII cells 36, but studies using isolated ATII cells have not demonstrated ICAM-1 expression 37. It is possible that this discrepancy may reflect differences in the methods used to detect ICAM-1, the use of different primary antibodies or loss of ICAM-1 during ATII cell isolation. ICAM-1 is upregulated by ATII cells, but not by ATI cells, in response to hyperoxia and various cytokines 36, 79, 80.
Receptor for advanced glycation end-products
The receptor for advanced glycation end-products (RAGE) is a multiligand-binding protein that binds advanced glycation end-products 81. Advanced glycation end-products are a heterogeneous group of molecules that accumulate in plasma and other body fluids as a consequence of age and certain diseases 81. The mRNA transcript for RAGE is highly expressed in lung tissue 81. Immmunoelectron microscopic studies demonstrate that RAGE is expressed on the basal surface of ATI cells in the lung 41, 42. However, mRNA transcripts for RAGE have also been detected in ATII cells in the lung 43. The role of RAGE in ATI cells is unknown.
Lectins
Finally, a group of lectins are able to distinguish between the carbohydrate moieties on the surface of ATI and ATII cells. Lectins such as Ricinus communis agglutinin 82, Lycopersicon esculentum agglutinin 83, Soybean agglutinin 84 and Bauhinia purpurea agglutinin 85 selectively bind to the surface of ATI cells.
Expression of alveolar epithelial type I cell-selective proteins following injury
Following injury, the amount of ATI-cell-selective proteins in lung tissue may change because of damage to ATI cells. However, ATI-cell-selective proteins may also change because they are also expressed on alveolar epithelial intermediate cell types (i.e. morphology-dependent changes in regulation) or because of changes in regulation by ATI cells (i.e. morphology-independent). The use of ATI cell-selective markers in the detection of ATI cell necrosis in both animal and human studies is reviewed below. Data from animal studies that suggest ATI-cell-selective proteins are upregulated by ATI cells in response to toxic agents or expressed by alveolar epithelial intermediate cells are also reviewed.
Use of rat type I cell 40-kDa protein/type I cell alpha protein and human type I cell 56-kDa protein to quantify ATI cell necrosis in ALI
Ultrastructural studies on injured lungs demonstrate that necrotic ATI cells are sloughed from their basement membranes into the airspaces 86–88. Various experimental studies demonstrate that the amount of RTI40/TIα protein in distal airway fluid or bronchoalveolar lavage fluid (BALF) can be used to quantify the extent of ATI cell necrosis (table 2⇓). Specifically, in a model of severe ALI, caused by Pseudomonas aeruginosa, RTI40/TIα protein levels are elevated 80-fold above control values in distal airway fluid 6 h after infection 89. Morphological analysis of infected lungs at this time point demonstrates widespread ATI cell necrosis and denuded alveolar basement membranes 89. In addition, the ability of the alveolar epithelium to clear fluid, a functional measurement of epithelial integrity, is completely abolished following P. aeruginosa-induced lung injury. In contrast, in a model of focal ATI cell necrosis, induced by nitrogen dioxide exposure, BALF concentrations of RTI40/TIα protein are elevated only two-fold above control values and alveolar fluid clearance is stimulated 90. BALF RTI40/TIα protein concentrations are also elevated, to varying degrees, in other acute injury models, including hyperoxia 91, Staphylococcus aureus-induced pneumonia 92, acid-instillation followed by ventilator-induced injury 96 and following intratracheal adminstration of bleomycin (table 2⇓) 94. In the rat model of acid-instillation, RTI40/TIα protein concentrations in oedema fluid and plasma were significantly reduced in animals ventilated at low tidal, or protective, volumes in comparison with those ventilated at higher tidal volumes (i.e. 3 versus 6 mL·kg body weight−1) 96. Low tidal volumes were also associated with better alveolar function as measured by the rate of alveolar fluid clearance 96. Combined, these studies suggest that ATI cell membrane proteins, such as RTI40/TIα protein, may be sensitive markers of lung injury resulting from a wide range of different causes. More specifically, however, these studies also suggest that the extent of ATI cell necrosis depends on the exact nature of the damaging agent.
Rat type I cell 40-kDa protein (RTI40) in rat lung injury models
ALI and ARDS are life-threatening conditions that may occur as a consequence of either direct (pneumonia and acid aspiration) or indirect (sepsis and trauma) injury to the lungs. As in experimental models of ALI, ultrastructural analysis of the lung in patients who have died due to ARDS demonstrates widespread epithelial damage, including ATI cell necrosis 97. In addition, clinical studies also demonstrate that preserved alveolar fluid clearance correlates with lower mortality in patients with ARDS 12. The concentration of various ATII-cell-selective proteins (e.g. surfactant protein A and D, and KL-6) in BALF and serum are altered in patients at risk of ARDS and with ARDS 98, 99. However, since the BALF concentration of these secreted proteins may well change independently of ATII cell necrosis, their usefulness in diagnostic assays may be limited.
At the present time, the antibodies that recognise the human homologue of RTI40 have not been used to quantify ATI cell injury in patients with ALI or ARDS. However, HTI56, a human ATI-cell-specific protein, has been measured in oedema fluid and plasma from patients with established ARDS. The concentration of HTI56 was significantly elevated in both oedema fluid and plasma from patients with ARDS in comparison to those from hydrostatic pulmonary oedema patients 100. These data suggest that ATI-cell-specific membrane proteins may be sensitive markers of alveolar epithelial injury in patients with, and at-risk of, ALI or ARDS.
Specific regulation of alveolar epithelial type I cell-selective proteins by toxic agents
Hyperoxia is frequently used as a model for the investigation of lung injury and repair. Hyperoxic exposure characteristically causes endothelial damage that is followed by ATI cell damage 101. Cao et al. 102 investigated the lung content of RTI40/TIα protein, AQP5 and caveolin-1 following hyperoxic exposure. Instead of a decrease in ATI-cell-selective protein levels (which might be expected if ATI cells were damaged), they found that RTI40/TIα protein and AQP5 were both upregulated by approximately five-fold, whereas caveolin-1 levels were unchanged. Immunohistochemical analysis, for RTI40/TIα protein, demonstrated that staining remained restricted to ATI cells and that there was no additional staining in other cell types. Moreover, using an alveolar epithelial cell line, Cao et al. 102 demonstrated that the transcriptional activity of the TIα promoter was directly regulated by hyperoxia. These data suggest that RTI40/TIα protein and AQP5 are regulated by ATI cells in response to hyperoxic stress.
In a mouse model of viral pneumonia, levels of AQP5 and its mRNA transcripts were both decreased 7 and 14 days following inoculation 103. Histochemical analysis of the alveolar wall suggested that this decrease in AQP5 expression was not due to ATI cell necrosis or increased numbers of ATII cells. Conversely, immunohistochemical analysis revealed that AQP5 staining was reduced throughout the lung, even in areas without overt inflammation. These data suggest that AQP5 is downregulated by ATI cells following viral pneumonia. AQP5 has also been reported to be downregulated in lung tissue following endotoxin-induced lung injury 104. The functional significance of AQP5 downregulation is not understood. Although viral and endotoxin-induced lung injury both cause extravascular fluid accumulation, studies in AQP5-deficient mice suggest that AQP5 does not play a role in fluid accumulation following injury 75.
Expression of alveolar epithelial type I cell-selective proteins in alveolar intermediate cells
Following injury to the lung, the alveolar walls are populated not only by ATI and ATII cells but potentially also by alveolar epithelial intermediate cells and airway or bone-marrow-derived progenitor cells. Very little is known about the proteins expressed by any of these cell types, even ATI and ATII cells, in the lungs following injury. However, data from a small number of in vivo studies suggest that alveolar epithelial intermediate cells may express certain ATI-cell-selective proteins.
Like NO2, ozone damages epithelial cells located at the junction between the terminal bronchioles and the first alveolar duct 105. Following ozone exposure, immunoelectron microscopy demonstrates that cytochrome P450 2B1 is located in ATI cells, as observed in noninjured lungs. However, P450 2B1 is also expressed in two other epithelial subtypes, which exhibit ultrastructural characteristics of both ATI and ATII cells 34. One of these epithelial cell types has microvilli, an extended and thick lateral cytoplasm, and large vacuoles that occasionally contain lamellar bodies. The other type exhibits a smooth surface and ATI-like lateral cytoplasmic extensions that sometimes show focal thickening 34.
Exogenous administration of keratinocyte growth factor stimulates ATII cell proliferation in vivo. Following keratinocyte growth factor administration, the number of ATII cells is maximal at 2–3 days; however, by 7 days, alveolar epithelial architecture appears normal 106. ATII cell numbers are reduced to control levels via both apoptosis and transdifferentiation to new ATI cells (interestingly, this process appears to be an example of ATII cell transdifferentiation that is not preceded by ATI cell injury). Prior to the return of the alveolar epithelium to normal, cells have been identified that exhibit both ATII and ATI characteristics. These characteristics include an elongated cytoplasm and expression of an ATI-cell-specific lectin, L. esculentum agglutinin, and an ATII-cell-selective cell protein, surfactant protein D 106.
Using confocal microscopy, the phenotype of the lung alveolar epithelium was investigated following intratracheal administration of S. aureus. A small population of cells that coexpress RTI40/TIα protein and an ATII-cell-selective antigen, MMC4, were identified 51, 107. RTI40/TIα protein/MMC4-positive cultured ATII cells have also been observed as they become ATI-like 108. These data suggest that RTI40/TIα protein/MMC4-positive cells are intermediates in the transition of ATII to new ATI cells (data not shown).
Changes in the surface expression of ICAM-1 have been investigated in a number of animal models using immunogold electron microscopic analysis. ICAM-1 levels on the surface of ATI cells decrease at cell boundaries following hyperoxia but do not change following pneumonia. However, ICAM-1 expression is dramatically increased on the surface of ATII cells following both pneumonia and hyperoxia 80, 109. Since ICAM-1 expression increases as ATII cells become ATI-cell-like in culture, the upregulation on ATII cells in vivo could be an early differentiation event 37. However, ICAM-1 expression is also upregulated in response to cytokines 79. Therefore, ICAM-1 expression may also be regulated by ATII cells independently of morphological changes associated with transdifferentiation to ATI cells.
To date, alveolar epithelial intermediate cells have been defined on the basis of their ultrastructural characteristics, and these characteristics appear to include at least two major intermediate cell types 20, 23, 34. The preliminary studies mentioned above suggest that alveolar epithelial intermediate cells may also be identifiable by investigation of the coexpression of ATI- and ATII-cell-selective markers. However, whether the identified coexpressing cells are ATII cells becoming ATI cells, or vice versa, remains unknown. Nor is it known whether different ultrastructural intermediates have different phenotypes. It is well known that the conditions under which isolated ATII cells are cultured influence whether or not they express ATII- or ATI-cell-selective proteins and mRNAs (reviewed in 25). Moreover, it has also been demonstrated that ATI-like cells can be made to re-express ATII-like phenotypic markers by varying the culture conditions, e.g. by going from submerged to air/liquid interface culture 110, 111. Studies designed to determine coexpression of ATI- and ATII-cell-selective markers in culture may help in the phenotyping of ATII-to-ATI intermediates, and in the determination of whether the same intermediates exist as ATI cells become ATII cells.
Alveolar epithelial type I cell-selective proteins and lung fibrosis
The term lung fibrosis encompasses a diverse group of lung diseases that share many clinical, radiographic and physiological characteristics. The most common and severe form of fibrosis is IPF. Morphologically, IPF is characterised by regions of fibrosis alternating with regions of normal lung with little evidence of inflammation 112. At the borders between fibrotic and normal lung are collections of fibroblasts/myofibroblasts, called fibrotic foci. These regions of active fibrosis are also characterised by the presence of different alveolar epithelial phenotypes 4. Immunohistochemical studies demonstrate that the alveolar epithelium in fibrotic regions expresses a number of profibrotic cytokines and growth factors 18. Based on these observations, it has been proposed that multiple cycles of alveolar injury and repair lead to aberrant alveolar epithelial cell populations that drive fibrosis 4. Although there is no disputing that the alveolar epithelium is not normal in IPF, the origin and specific functions of the different epithelial phenotypes remain unknown. It has been proposed that some of the distinct alveolar epithelial cells in IPF are alveolar epithelial intermediate cells 4, 113.
Bleomycin is commonly used to induce pulmonary fibrosis in experimental animals, although, unlike IPF, bleomycin-induced fibrosis is classically associated with inflammation. Four weeks after bleomycin treatment, regions of the alveolar barrier stain positively for some ATI-cell-specific lectins (e.g. B. purpurea agglutinin) but demonstrate focal loss of RTI40/TIα protein 94. Other investigations into lung fibrosis also report the loss of ATI-cell-associated proteins, including caveolin-1, RTI40/TIα protein, AQP5 and RAGE 30, 95, 114. More studies are required in order to fully understand to what extent the phenotype of ATI cells change in fibrosis or whether the loss of ATI-cell-selective proteins reflects a decrease in the number of ATI cells. Data from knockout mice, and in particular TIα-deficient mice (see earlier) 63, suggest that TIα (or ATI cells) may play a role in the regulation of distal airway cell proliferation. Therefore, it is possible that loss of ATI cells in fibrosis not only leads to a decrease in surface area for gas exchange but may also contribute to the increased proliferation of other distal airway cells.
Conclusions
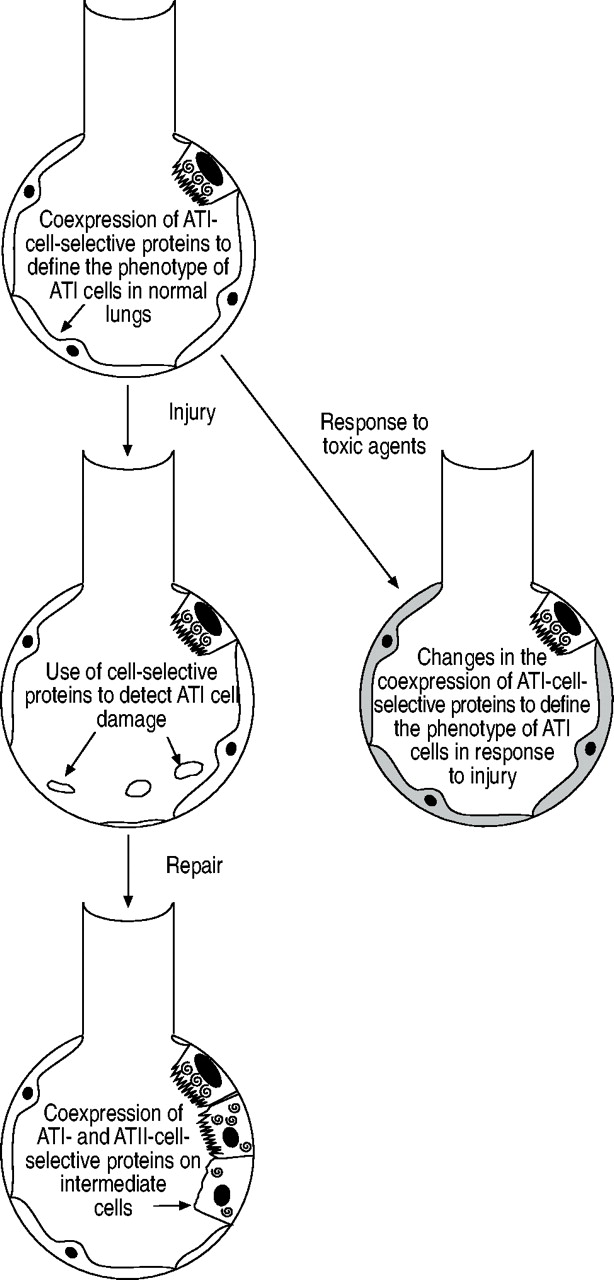
The present article has reviewed the use of cell-selective proteins as markers for the investigation of ATI cell injury and repair following injury (fig. 3⇓). ATI-cell-selective proteins such as RTI40/TIα protein are useful for quantifying the extent of ATI cell necrosis in animal models of lung injury, and human homologues of RTI40/TIα protein may form the basis of diagnostic assays in conditions such as ALI or ARDS. The coexpression of ATI-cell-selective markers, such as RTI40/TIα protein, AQP5, caveolin-1 and 2, and cytochrome P450 2B1, may help identify ATI cells in immunohistochemistry- and immunofluorescence-based studies and facilitate investigation into ATI cell numbers at active sites of injury and repair. The coexpression of ATI cell-selective proteins with ATII- selective proteins may help identify alveolar epithelial intermediate cells and their role in lung repair and fibrosis.

{kind=link}
{kind=link}
{kind=link}
Potential applications of cell-selective markers in the investigation of alveolar epithelial type I cell injury and repair. ATI: alveolar epithelial type I.
A better understanding of the regulation of alveolar epithelial type I cell numbers, and the function of proteins selective for alveolar epithelial type I cells, will lead to a greater understanding of the role of alveolar epithelial type I cells in both normal and diseased lungs.
Acknowledgments
The authors kindly thank L. Dobbs (University of California at San Francisco, San Francisco, CA, USA) for the monoclonal antibody directed against rat type I cell 40-kDa protein. They also thank J.V. Gray (University of Glasgow, Glasgow, UK) and S. McKechnie, L. Franklin and K. Tyrrell (all University of Edinburgh, Edinburgh, UK) for comments on the manuscript and G. Clegg (University of Edinburgh, Edinburgh, UK) for assistance in preparing figure 1⇑.
- Received August 21, 2003.
- Accepted April 29, 2004.
- © ERS Journals Ltd
References









