





Abstract
To study the role of small airways in the early allergic response (EAR), the method of human precision-cut lung slices (PCLS) was developed and used to examine the bronchoconstriction elicited by passive sensitisation and allergen provocation.
Viable human PCLS of 250-µm thickness containing airways <1.5 mm in outer diameter were prepared from lung lobes obtained from lung resection and taken into culture. According to the low release of lactate dehydrogenase and the constant ciliary beat frequency, human PCLS were viable for at least 3 days.
Following overnight passive sensitisation with serum from allergic individuals, administration of grass-pollen extract or activating immunoglobulin E antibody resulted in immediate airway contraction that was quantified by videomicroscopy. The extent of the EAR increased with decreasing airway size (outer airway diameter), with the strongest response occurring in the terminal bronchioles.
Histamine receptor antagonism was ineffective, and leukotriene or thromboxane receptor antagonism attenuated the early allergic response only in some cases. However, simultaneous blockade of leukotriene and thromboxane receptors almost completely prevented the early allergic response in the precision-cut lung slices from all individuals, suggesting such a dual treatment as a potential future asthma therapy.
This study was supported by the Deutsche Forschungsgemeinschaft grant DFG Uh 88/3-1 to S. Uhlig.
Bronchoconstriction and airway hyperreactivity are major symptoms of asthma. The role of small airways (<2 mm in diameter) in the disease process of bronchial asthma is still unresolved, but awareness of their importance is growing 1–3. On the basis of measurements with the retrograde catheter technique in animals, Macklem and Mead 4 determined that the small airways contribute <10% of the total airway resistance and hence termed them the “quiet zone”. More recently, however, using more sophisticated techniques, van Brabandt et al. 5 suggested that peripheral resistance contributes between 50–90% of the total resistance in normal lungs and in the lungs of patients with chronic obstructive lung disease. Given that such a large fraction of airway resistance resides in the peripheral airways, it is not surprising that a number of clinical studies suggested peripheral airways as the predominant site of airflow obstruction in asthmatics 6–9. Moreover, substantial airway inflammation is present in the peripheral airways of asthmatic patients 3, 10 and computational analyses based on quantitative histology led to the conclusion that the peripheral airways account for the majority of the airway hyperresponsiveness among these persons 11, 12. Because airway inflammation and hyperresponsiveness are thought to be related to previous mast-cell degranulations during the early allergic response (EAR), responsiveness to allergens may, at least, partly explain these pathological alterations of small airway function and structure. However, with one exception, the contribution of small airways to the EAR has not been investigated in humans. Using sensitised bronchi, Ellis et al. 13 showed that airways down to 0.5 mm in diameter are capable of an EAR. However, their method did not permit the authors to investigate airways of <0.5 mm and to establish a correlation between airway size and the strength of the EAR.
In the past, the difficulty to perform functional studies with peripheral areas of the lung has hindered progress in this area of research. New insight into the small airways has become possible with the model of precision-cut lung slices (PCLS), which allows the analysis of the responses of airways down to the terminal bronchioles by quantitative videomicroscopy 14. With this method, lung tissue, including airways, is cut into thin viable PCLS of ∼250 µm thickness that are taken into culture. Functional parameters such as bronchoconstriction can be studied under a microscope and digital-imaging techniques make it possible to visualise and quantify contraction of airways. In contrast to conventional in-vitro methods of measuring bronchoconstriction, airway contraction in vivo is neither isometric nor isotonic, because in vivo the load that the muscle has to overcome increases as is shortens. Videomicroscopy of airways allows the determination of airway contraction and dilation under quasi-auxotonic conditions 15. Another advantage of this method is that thetotal incubation volume is rather small (0.5–1.0 mL) sothat the dilution of endogenously produced mediators isminimised.
Using rat PCLS, it has been shown previously that the EAR increases with decreasing airway size, being maximal at the terminal bronchioles 16. However, asthma is difficult to model in experimental animals, for instance, serotonin, an important mast cell mediator of the EAR in rats 16, is almost absent from human mast cells 17. The present study had three goals. First, to establish the method of PCLS for the measurement of bronchoconstriction and ciliary beat frequency in human lung tissue. Second, to use passively sensitised PCLS to investigate the significance of human small airways in the pathophysiology of asthma. Third, to use this novel model for pharmacological investigations of the EAR in human airways.
Methods
Human precision-cut lung slices
Human lung slices were prepared from patients undergoing surgery for cancer, most of them smokers. Carcinomas were later identified in the division of pathology as nonsmall cell carcinomas including squamous cell carcinoma and adenocarcinoma. Macroscopically healthy lung tissue at distance to the tumour was used to produce PCLS. The experiments were approved by the local ethics committee.
Macroscopically healthy areas were filled via the main bronchus with 500–800 mL low melting point agarose solution (final agarose concentration 1.5%), and put on ice until the agarose had solidified. The used volume of agarose corresponds approximately to the total lung capacity (TLC) of this part of the lung 18. An area distal to the tumour was then separated from the rest of the lobe, which remained with the pathologist for further examination. The lung tissue was cut further into 5–10 mm thick slabs from which cores along the airways were made with a coring tool. These cores, with a diameter of 0.9 cm, were cut into ∼250 µm thick slices by a Krumdieck tissue slicer (Alabama Research and Development, Munford, AL, USA). Subsequently, the PCLS were incubated in minimal essential medium (MEM; GIBCO Life Technologies, Eggenstein, Germany) and medium was changed every 30 min for the first 2 h and then every hour for the following 2 h to remove the agarose and mediators from the airways. Lung slices were cultured in medium based on MEM. The 10× concentrated MEM solution (w/o NaHCO3, w/o phenol red) was supplemented with amino acid solution (GIBCO), 200 nM sodium pyruvate (GIBCO), vitamin solution (GIBCO), glucose (final concentration 2.2 mM), N-2-hydroxyethylpiperazine-N-2-ethanesulphonic acid (HEPES) (4.6 mM) and NaHCO3 (5.2 mM). pH was adjusted to 7.4 before sterile filtration. Penicillin and streptomycin (GIBCO) were added to the MEM solution at a final concentration of 100 U·mL−1 and 100 µg·mL−1, respectively. For instillation of agarose, double-concentrated medium was mixed with the same volume of low melting point agarose solution at 37°C. Slicing was performed in MEM buffer. MEM powder (without NaHCO3, with phenol red) with Earl's salts was supplemented with 6 g·L−1 HEPES and 2.2 g·L−1 NaHCO3. The pH was adjusted to 7.4.
Imaging and analysis
The airways were imaged and digitised with a digital video camera as described earlier 16. Airway area before addition of the allergen was defined as 100%. Bronchoconstriction was expressed as the percentage decrease in airway area in comparison with the control airway area.
Assessment of airway precontraction
PCLS were cultured in MEM and the medium changedroutinely to remove agarose and avoid effects of endogenously released mediators. Precontracted airways show a distinct folding of the airway epithelium and the portion of airway smooth muscle is >50% of the outer airway area. From these observations, an experienced experimenter may estimate the degree of precontraction of airways. To verify this procedure, airways of 14 PCLS, which were not used for further experiments, were first assessed by the experimenter before they were treated with 10 µM isoproterenol (Sigma, Taufkirchen, Germany) to obtain maximal dilation. The airway area and diameter was imaged before and after addition of the bronchodilator. Two of 14 PCLS were categorised as precontracted by the experimenter. Treatment with isoproterenol increased the median airway area to 117%. In contrast, the two PCLS, categorised as precontracted, responded to isoproterenol by dilating to 170 and 345%, respectively. From these findings and from other observations made during this study, the authors concluded that ∼10% of all airways were precontracted. Such airways were excluded from this study.
Viability
Viability of human PCLS was determined by measuring lactate dehydrogenase (LDH) leakage into the incubation medium and determining ciliary beat frequency over a time period of 3 days.
Lactate dehydrogenase leakage
Three PCLS per well were incubated in a 24-well plate and covered with 1 mL medium. At 0, 12, 14, 36, 48, and 72 h slices were removed and put into 1 mL 0.2% Triton X-100 and kept on ice until further processing. The supernatant was also removed and put on ice. PCLS samples were prepared for LDH measurement by tissuehomogenisation in a Polytron tissue homogeniser (Kinematica AG, Littau, Switzerland). All samples, including the supernatant, were then centrifuged for 20 min at 11,000×g at 4°C to remove cell debris. Viability was expressed as the percentage LDH activity (Dimension pan; Dade Behring, Schwalbach, Germany) in the supernatant of the total LDH activity.
Ciliary beat frequency
Beating of cilia along the epithelial border of the airways was recorded with a fast camera (Visicam 640; Visitron Systems, Puchheim, Germany) recording 62 images per second at a magnification of 640. Along the epithelial border virtual lines were drawn representing 256 pixels. For each pixel, the change in light intensity was recorded and plotted against time. Ciliary beat frequency was determined by Fourier analysis. In each image measurements were obtained at three different sites of the luminal border. Such measurements were performed on three PCLS per patient. The data from each slice were averaged and the mean of the average reported.
Passive sensitisation and induction of the early allergic response
Human PCLS were treated overnight with 1% serum from patients with allergic rhinitis. Serum from three different patients with low specific immunoglobulin (Ig)E (2.8 U·mL−1), medium specific IgE (50 U·mL−1) and high specific IgE (420 U·mL−1) aged 35, 50 and 28 yrs, respectively, was used. The specific IgE against timothy grass (Phleum pratense) pollen was quantified by fluorescent enzyme immunoassay (CAP-FEIA; Amersham Pharmacia, Freiburg, Germany). In addition, human PCLS were also treated with an activating goat antihuman IgE (100 ng·mL−1; Biosource, Nivelles, Belgium).
Single PCLS were incubated with 1 mL of serum-containing MEM at 37°C and 5% carbon dioxide in an incubator overnight (∼15 h). The following day, the PCLS received fresh medium free of antibiotics and serum. Single slices were moved from the incubator onto a fresh 24-well plate, positioned within the well and then put under a microscope (Inverted microscopes Leica DMIRB and DMIL; Zeiss, Oberkochen, Germany). A control image was taken before addition of the allergen (5 µg·mL−1 Phleum pollen extract), which was administered in prewarmed (37°C) MEM. Image acquisition was started after addition of the solution. Images were made every 30 s for 10 min.
Pharmacological intervention studies
To investigate the mediators of the EAR, PCLS were preincubated for 10 min with the leukotriene (LT)-receptor antagonist montelukast (MSD Sharp & Dohme GmbH, Haar, Germany), the thromboxane-receptor antagonist SQ 29548 (SPI Bio, Paris, France) and the histamine (H)1-receptor antagonist triprolidine, either alone or in combination prior to challenge with the allergen.
Statistics
The correlation analysis was performed by calculating the Spearman rank coefficient. The α-error due to multiple comparisons was controlled by using the Bonferroni-Holm procedure.
Results
The preparation of human precision-cut lung slices
Preparation of human lung tissue yielded up to 30 slices from one resection with airways ranging from generation 12 (∼1.5 mm in diameter 18) down to the terminal bronchioles (<0.6 mm in diameter). Viability of human PCLS was determined by measuring LDH leakage into the incubation medium and ciliary beat frequency over a time period of 3 days. LDH release remained <10% for the first 24 h and did not exceed 15% within 72 h (fig. 1⇓). In membranous airways of human PCLS, ciliary beat frequency ranged between 5–12 Hz at 37°C, exhibiting only slight changes within 72 h (table 1⇓). These findings demonstrate that human PCLS are viable for at least 3 days.

Leakage of lactate dehydrogenase (LDH) from human precision-cut lung slices over time. Lung slices were incubated in 24-well plates for up to 72 h in an incubator at 37°C and 5% carbon dioxide. Values shown are the mean±sd from three independent experiments.
Ciliary beat frequency in human precision-cut lung slices (PCLS) from individual lung donors determined on 3 consecutive days after preparation of the PCLS
To further establish this model, the authors examined the responses of PCLS to acetylcholine and its stable analogue methacholine. Airways in human PCLS responded to acetylcholine by an immediate airway contraction that was reversible within 30 min (fig. 2a⇓). The median effective extracellular concentration (EC50) for methacholine was 0.4 µM (fig. 2b⇓).

a) Time-dependency of acetylcholine (Ach) on airways of human precision-cut lung slices (PCLS) showing the contraction and dilation of the airways after exposure to Ach (10−6M). Values are the mean±sd from PCLS of three different patients. b) Concentration/response curve of methacholine (Mch) in small human PCLS. Mch was used for cumulative measurement at concentrations from 10−9–10−4 M. Values shown are the mean±sem from four to seven different patients. The medium airway size was 1.18 mm in outer airway diameter.
The early allergic response
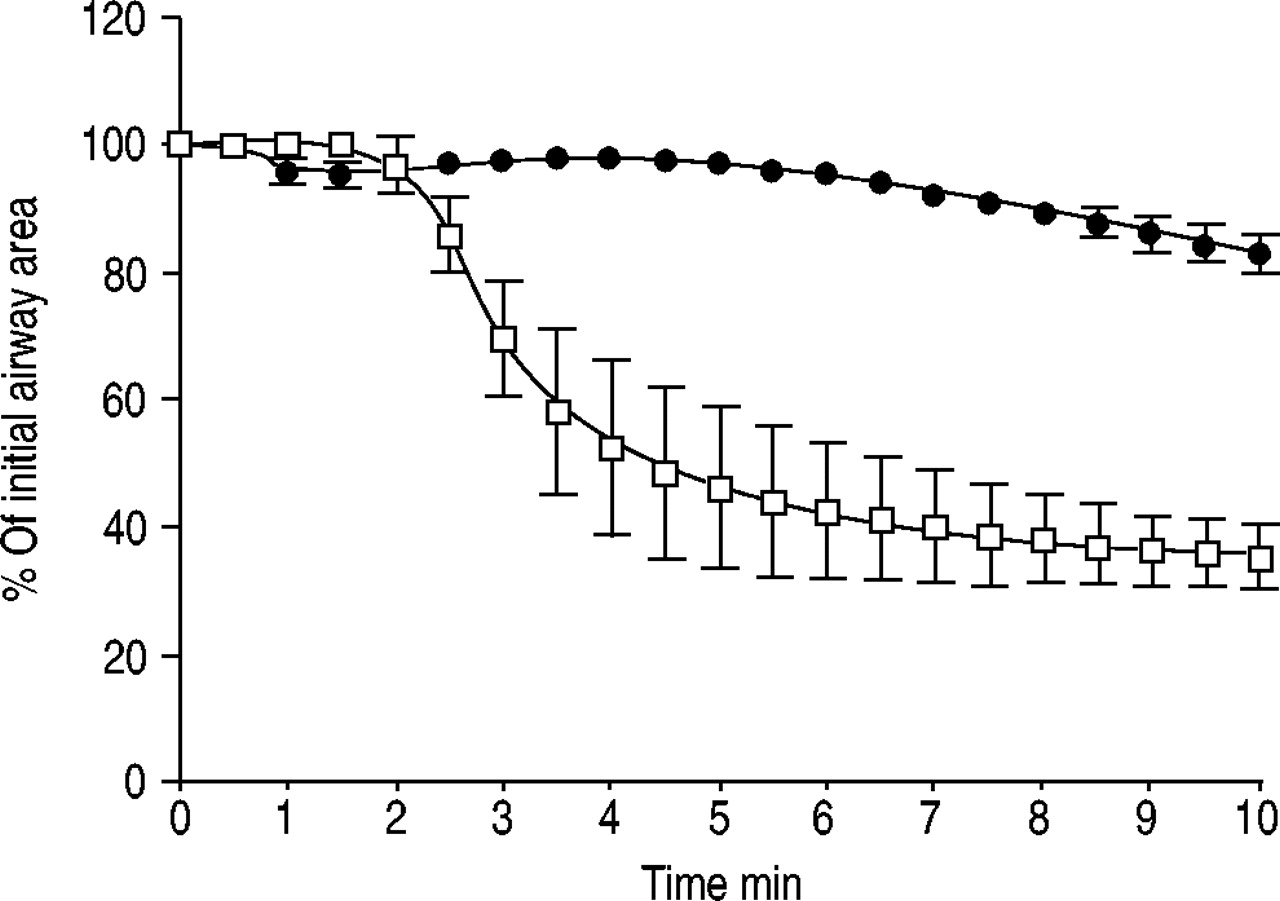
Human PCLS prepared from nonatopic patients were incubated overnight with sera from three different nonatopic patients containing high, medium and low concentratrions of IgE, specific for the pollen of timothy grass (P. pratense). If slices were incubated with serum containing high or medium levels of specific IgE, airways contracted within 2 min of antigen challenge (figs 3 and 4⇓⇓). If, however, slices were incubated with serum from a nonatopic donor with a low level of specific IgE (2.8 U·mL−1), addition of pollen extract had no effect (fig. 4⇓). Furthermore, in nonsensitised PCLS, pollen extract caused no bronchoconstriction and a nonspecific antigen, such as ovalbumin, failed to induce bronchoconstriction in sensitised PCLS (data not shown). In contrast, incubation with an IgE-activating antibody caused contraction in sensitised PCLS (fig. 5⇓). This indicates that the EAR in human PCLS is dependent on IgE.

Images of a lung slice showing a single airway a) before and b) 10 min after administration of 5 µg·mL−1 pollen extract. The human precision-cut lung slices were treated overnight with 1% serum from an atopic donor with a specific immunoglobulin-E titre (50 U·mL−1) against timothy grass (Phleum pratense) pollen (passively sensitised). A: airway lumen. Internal scale bar=1 mm.

Time course of airway contraction in sensitised precision-cut lung slices (PCLS) after addition of 5 µg·mL−1 pollen extract. Human PCLS were treated overnight with 1% serum from three atopic donors with low specific immunoglobulin (Ig)E (2.8 U·mL−1, n=4; ▪), medium IgE (50 U·mL−1, n=16; □) and high IgE (450 U·mL−1, n=4; •) titres against timothy grass (Phleum pratense). Values shown are the mean±sem.

Time course of immunoglobulin (Ig)E-induced bronchoconstriction in sensitised and unsensitised precision-cut lung slices (PCLS). Human PCLS were incubated overnight with either a 1% serum high IgE titre (□) or with minimum essential medium only (•). Subsequently, the PCLS were treated with an activating anti-IgE antibody (100 ng·mL−1). Values shown are the mean±sem from PCLS obtained from three different lungs.
Effect of airway size on the early allergic response
In PCLS, airway size can be measured and related to the responsiveness of each individual airway. Because human airways may have an inherent tone, which may affect the inner airway diameter, the authors used the outer airway diameter as a measure of airway size. In addition, airways that appeared to be precontracted were excluded from the analysis. Figure 6⇓ shows a scatter diagram of the outer airway diameter plotted against the maximum degree of airway contraction observed within 10 min of antigen challenge. A correlation was found between airway size and the extent of contraction (r=0.35, p=0.0035), in that the strength of the antigen-induced contraction increased from larger airways to the terminal bronchioles.

Correlation diagram of the extent of airway contraction induced by pollen extract in sensitised human precision-cut lung slices (PCLS) plotted against airway size (outer airway area). PCLS with airways of different sizes were passively sensitised overnight and subsequently challenged with 5 µg·mL−1 pollen extract the next day. Each value represents the maximum contraction of a single airway with the given airway size. The top x-axis indicates the corresponding outer airway generation according to Yeh and Schum 18. A total of 70 single lung slices from 25 nonatopic individuals (17 male, eight female, aged 42–77 yrs) were investigated. Data were analysed by calculating the Pearson correlation coefficient (r=0.35, p=0.0035). #: terminal bronchioles.
Pharmacological characterisation of the early allergic response in human precision-cut lung slices
To identify the mediators responsible for inducing bronchoconstriction during the EAR, slices were preincubated withdifferent inhibitors, either alone or in combination. Preincubation of passively sensitised PCLS with the serotonin-receptor antagonist ketanserin (100 µM) before challenge did not affect the EAR (data not shown).
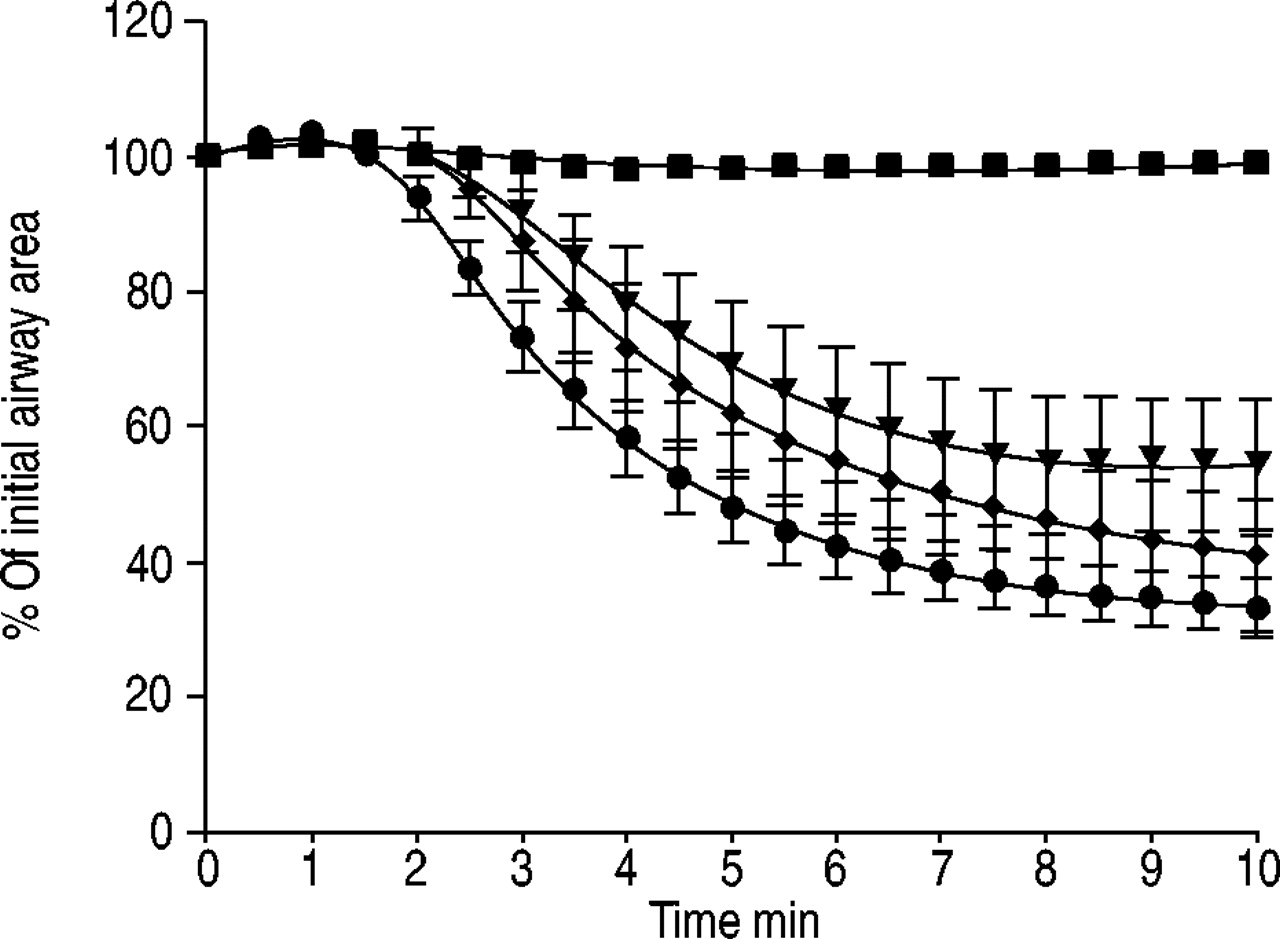
The individual responses of the PCLS from each of the 12 lung donors to pretreatment with the thromboxane-receptor antagonist SQ 29548 (10 µM), the H1-receptor antagonist triprolidine (5 µM) and/or the LTD4-receptor antagonist montelukast (10 µM) are shown in figure 7⇓. All agents were used at a concentration at least 1,000-fold above their median inhibitory concentration values 19, 20. Bronchoconstriction was reduced in response to antigen challenge in three of 12 independent individual's PCLS pretreated with SQ 29548 (fig. 7⇓) and in four of 12 pretreated with montelukast, whereas triprolidine appeared to be largely ineffective. Combination of triprolidine with either SQ 29548 or montelukast failed to attenuate the bronchoconstriction to allergen in PCLS. Of note, histamine-induced bronchoconstriction in human PCLS appeared normal with a maximal contraction of up to 40% ofthe initial airway area and an EC50 of 3 µM (−log10 EC50=5.5±0.14, n=5). The combination of SQ 29548 and montelukast largely prevented the EAR in all cases (12 of 12). The kinetics of the allergen-induced bronchoconstriction were comparable in slices treated with SQ 29548 or montelukast (fig. 8⇓).

Effect of cysteinyl-leukotriene-, histamine 1-, and/or thromboxane-receptor blockade on the early allergic response in human precision-cut lung slices (PCLS). PCLS were passively sensitised overnight and subsequently challenged with 5 µg·mL−1 pollen extract the next day. Ten minutes before allergen challenge, airways were pretreated with 10 µM montelukast (Monte), 10 µM SQ 29548 (SQ) or 5 µM triprolidine (Tripro). Shown are a) the combination of Monte and SQ, b) the combination of Tripro and SQ and c) the combination of Monte and Tripro. The data represent slices from 13 different nonatopic individuals (eight females, five males, mean age 58.6 yrs). Medians are shown as horizontal lines. **: p<0.01 versus pollen extract (PE) alone.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Time course of pollen extract-induced bronchoconstriction. Sensitised human precision-cut lung slices were treated with grass (Phleum pratense) pollen extract (•, n=22) in the presence of montelukast (▾, n=11), SQ 29465 (♦, n=15) or the combination of the two inhibitors (▪, n=7). Values shown are the mean±sem.
As experimental animal studies have suggested that LTs exhibit their action via the formation of thromboxane 21, lung slices were preincubated with SQ 29548 to block thromboxane receptors and subsequently challenged with LTC4 or LTD4. Administration of LTC4 and LTD4 induced bronchoconstriction, despite pretreatment with SQ 29548 (table 2⇓). The authors therefore conclude that in human lungs, LTs exert bronchoconstriction independently from thromboxane.
Leukotriene (LT)-induced bronchoconstriction in human precision-cut lung slices (PCLS)
Discussion
Asthma is characterised by airway obstruction, inflammation of the airway mucosa, and airway hyperresponsiveness. The initial step of the allergic airway obstruction is the EAR. In this study, the focus was placed on the response in small airways. The data suggest that the EAR can take place in small human airways and may be treated effectively by dual administration of LT- and thromboxane-receptor antagonists.
The model of precision-cut lung slices
Because of the possibility to examine airways of all airway generations with diameters <5 mm, PCLS are suited to investigate the responsiveness of individual airways in relation to their size. To this end, single slices are monitored by videomicroscopy, and airway size, airway contraction, vessel contraction and ciliary beat frequency are determined by digital-imaging techniques. Taking advantage of this method, the authors have recently defined the responsiveness of small airways to methacholine 14, thromboxane 22, endothelin 22 and allergen 16 in rats. Until now, human PCLS have been used in toxicological studies 23, but not to investigate the responses of single human airways. As judged from the release of LDH and the constant ciliary beat frequency, human PCLS were viable for ≥72 h. The present observations match those made previously, which indicated that PCLS are viable for ≥1–3 days 14, 23.
There are several potential problems in the use of PCLS to measure small airway responses. 1) Are airways in PCLS precontracted and how would this affect the measurements? 2) Is there load on airway smooth muscle? 3) Does instillation of agarose and inflation to TLC change airway responses? 4) Is the use of airway area instead of the diameter adequate for the analysis of bronchoconstriction?
In response to these questions: 1) all PCLS with strong visible precontraction were excluded from this study. Thus, although it cannot be completely ruled out that some airways were precontracted, it may be assumed that the extent of precontraction in the PCLS that were used in this study was only small. Furthermore, to exclude any effect of precontraction, in the correlation analysis the authors used the outer rather than the inner airway diameter. Finally, similar results to those reported here were obtained in airways of rats 16, which lack an inherent airway tone 14. 2) Lung recoil resides primarily in the lung parenchyma which is under some tension even at low lung volumes 24. As small airways are directly connected to the parenchyma this is expected to put some load on the airway smooth muscle in small airways with an intact surrounding microanatomy as in PCLS. However, at present there is no data on whether airway smooth muscle in PCLS is loaded or not. Nonetheless, given the fact that the microanatomy is maintained and that the PCLS reopen once the stimulus is removed (fig. 2⇑) 16, the authors believe that airway smooth muscle is loaded at least to some extent in PCLS. Certainly, load builds up once the airways start to contract and the surrounding tissue is distorted. The load that builds up during contraction may be enhanced by the fact that the lung tissue is held in place by the weight of a platinum wire as described 14. Furthermore, if lack of load would significantly alter airway responses in PCLS, the airways in PCLS would be expected to contract more readily than in the intact lung. However, as the authors have shown previously, for various mediators, the median effective concentrations required for airway contraction and increases in airway resistance are the same in PCLS and in intact lungs, respectively 25. 3) The preparation of PCLS requires that lungs be filled with high volumes of agarose approaching TLC 18. Adler et al. 26 investigated the elasticity of air-filled lungs in contrast to agarose-filled lungs and found no difference in the elasticity of the parenchyma in their preparation. Another concern is that the high pulmonary pressures of up to 30 cmH2O, used for filling the lungs with agarose, may alter recoil and tethering forces as well as the parenchyma-airway interdependence. Dandurand et al. 27 investigated the effect of different agarose filling volumes on airway area and methacholine responsiveness in razor blade-cut rat lung slices. They observed that increasing lung volume resulted in increased airway area, but had no effect on maximal airway narrowing (if comparing filling with 50 or 75–100% TLC, although a small effect between 25–100% TLC was reported) or the EC50. Thus, filling the lungs at TLC appears to increase the size of airways, but not their responsiveness. In addition, because these authors did not report the outer airway diameter, it cannot be excluded that at the lower filling volumes the airways were not fully recruited. Finally, these authors did not find any differences related to airway size. 4) With respect to the calculation of airway responses, it may be asked whether utilisation of the airway area will not exaggerate any possible role of small airways, because any change in area corresponds at least to a square change in radius. However, according to Poiseulle's law the determinant of airflow is the fourth power of the radius, suggesting that airway area is a better approximation than airway radius. This is also supported by the excellent correlation between EC50 values obtained from area measurements in PCLS and from measurements of pulmonary resistance in isolated perfused lungs 25. Moreover, the relative changes remain identical regardless of whether area or radius is used. For instance, if in figure 6⇑ the correlation coefficient is calculated by using outer airway diameter instead of area, the same value for the nonparametric Spearman correlation coefficient (0.32, p=0.0075) is obtained, and only slightly different values if Pearson's (parametric) correlation coefficient is calculated (0.35, p=0.0035 based on diameter versus 0.34, p=0.0051 based on area).
Taken together, measurement of airway area in PCLS appears to be a reliable tool to study small airway responses. EC50 values obtained with PCLS or with conventional methods are in the same order. For example, the EC50 of 3 µM for histamine-induced bronchoconstriction in human PCLS matches that in human tracheal preparations 28. The EC50 value for methacholine-induced bronchoconstriction in human PCLS was 0.4 µM compared with 1 µM in isolated human bronchi 29. This difference may be explained by the observation from rat PCLS that smaller airways are more sensitive to methacholine 14.
Airway size and the early allergic response
Small airways have been defined as those with a diameter of <2 mm 1, 2. As a result of their relative inaccessibility, their role in human asthma has only been poorly defined. In the present study, the authors demonstrate that PCLS offer a novel approach to study bronchoconstriction in small human airways. While these findings clearly demonstrate that sensitised small airways in PCLS respond to allergen, it is difficult to predict whether these observations also reflect the ability of small airways in intact human lungs to respond to inhaled stimuli. As noted above, contractile responses of airways of any size are a complex process depending on the forces exerted by the surrounding tissue and on the load on the airway smooth muscle. Nevertheless, these findings are in line with and extend those clinical and experimental findings summarised above, which all suggest an important role of small airways in asthma. In addition, at least in the case of thromboxane, the greater sensitivity of small airways within PCLS 22 is confirmed by histological analysis of intact lungs 30. Finally, some clinical studies have shown superior efficacy on asthma symptoms of aerosols with improved delivery to the lung periphery 31–33.
Extrapolation to the situation of human asthmatics is also complicated by the poor understanding of inhaled allergen deposition. Upon inhalation the relatively large pollen particles (20–30 µm) will most probably not immediately reach the lung periphery, but it has been suggested that they may agglomerate in the small airways independently from daily pollen exposure, as a result of the gradual cumulative effect of deposition of small amounts of allergen in the lower airways 34. More recently, however, it was demonstrated that grass pollen allergens might form aerosols with particle sizes between 0.12–4.67 µm 35, small enough to reach the small airways. It also seems possible that therapeutic strategies, which preferentially open larger airways, may help to channel allergens to the smaller airways, thereby shifting symptoms towards the lung periphery.
With all these reservations in mind, the regression analysis shown in figure 4⇑ nonetheless indicates that the EAR became stronger with decreasing airway size, as the slope of the regression line between airway size and the extent of bronchoconstriction was significantly different from zero, although this effect was less pronounced in human compared with rat PCLS 16. Why small airways, at least in PCLS, exhibit a stronger response to allergen is unknown. They appear not to be more sensitive in general, as is exemplified by endothelin-1, to which all airway generations respond similarly 22. A possible explanation would be a heterogeneous distribution of mast cells, which are mainly responsible for the mediator release during the EAR 36, 37. In fact, based on lung wet weight, allergen-challenged peripheral lung tissue releases more prostanoids, LTs and histamine than bronchial tissue 13, 38. Alternatively, a larger percentage of unoccupied IgE receptors on mast cells in peripheral airways than central airways might lead to a facilitated degranulation of these mast cells 13. A third possibility is that cysteinyl (Cys)LT1 and thromboxane prostanoid (TP) receptors, both of which have been identified in great numbers in the human lung 39–41, are heterogeneously distributed throughout the bronchial tree. Finally, structural differences between large bronchi and small bronchioles, such as the relatively greater muscular thickness in peripheral airways might account for differences in airway reactivity 42, 43.
Hence, smooth muscle shortening may result in greater narrowing of smaller airways 44. As a note of caution, most of the lung tissue used for the preparation of PCLS was obtained from smokers. Since smoking of >10 packs per year can lead to peripheral airway inflammation and increased airway responsiveness 45, the authors cannot exclude effects on smooth muscle shortening by smoking habits.
Mediators of the early allergic response
The authors have no direct evidence for the presence of mast cells in PCLS, but it is generally accepted that the EAR is caused by IgE-dependent mast cell activation 17. While the role of mast cells was not addressed in the present study, the essential role of IgE for the sensitisation of lung tissue was confirmed by the dependency on IgE in the serum (fig. 4⇑) and by the bronchoconstriction induced by an IgE-activating antibody (fig. 5⇑). Upon triggering, pulmonary human mast cells release a variety of different mediators, among them bronchoconstrictors such as histamine, LTs, prostaglandin D2 and thromboxane 17. These mediators cause airway contraction by binding to specific receptors on airway smooth muscle, i.e. histamine to H1 receptors, LTs to CysLT receptors and both prostaglandin D2 and thromboxane to TP receptors 46, 47. Previously, Ellis et al. 13 demonstrated in isolated human bronchi that the combined triple inhibition of prostanoids, LTs and histamine largely prevented allergen-induced bronchoconstriction in small and large human airways. However, they made no attempt to further identify the specific roles of these mediators. In the present study, the authors extended these observations by investigating the effect of all possible one-way and two-way combinations of H1-, CysLT- and TP-receptor antagonists. They found that the H1-receptor antagonist triprolidine was ineffective, and CysLT- or TP-receptor blockade was effective only in some cases. These findings are in line with no proven and documented benefit of antihistamines in asthma 48, and with the clinical observations of CysLT1- and TP-receptor antagonists being effective only in subsets of patients 46–51. Notably, however, simultaneous blockade of CysLT- and TP-receptors provided nearly complete protection from antigen-induced airway contraction in PCLS from all individuals. The mechanistic basis for this synergism is unknown, but unlike in guinea-pigs 52 and rats 53, in humans, LTs do not exhibit their action via the secondary release of thromboxane 54 (table 2⇑). These findings suggest that in humans the EAR is mediated by a synergistic effect of TP-receptor agonists and peptido-LTs, at least in the small airways. To the best of the authors' knowledge, with the exception of a recent single case study 55, in humans the cooperative effects between a CysLT-receptor antagonist and a TP-receptor antagonist have not been described before.
The authors conclude that precision-cut lung slices offer a useful way to study physiological responses in human lung tissue. Using this novel method, they have shown that an early allergic response is principally possible even in very small human airways. Acknowledging some reservations as to possible differences between small airways in situ and in precision-cut lung slices, their findings further suggest that the early allergic response may become stronger with decreasing airway size. Finally, these data indicate that simultaneous targeting of leukotriene and thromboxane receptors in small airways may be considered as a novel treatment in asthma.
Acknowledgments
The perfect technical assistance of C. Rodde and D. Buchholz is gratefully acknowledged.
- Received April 3, 2002.
- Accepted February 12, 2003.
- © ERS Journals Ltd
References









