





Abstract
Endothelin-1 (ET-1) has been shown to have a constrictor effect on the airways and parenchyma; however, the roles of the ETA and ETB receptors in the ET-1-induced changes in the airway and tissue compartments have not been fully explored.
Low-frequency pulmonary impedance (ZL) was measured in anaesthetized, paralysed, open-chest guinea-pigs. ZL spectra were fitted by a model to estimate airway resistance (Raw) and inertance (Iaw), and coefficients of tissue damping (G) and elastance (H), and hysteresivity (η=G/H).
Two successive doses of ET-1 (0.05 and 0.2 nmol·kg−1) each evoked significant dose-related increases in Raw, G, H and η. Pretreatment with 20 nmol·kg−1 BQ-610 (a highly selective ETA receptor antagonist) resulted in a significantly decreased elevation only in H after the lower dose of ET-1. However, all parameters changed significantly less on the administration of ET-1 after pretreatment with 80 nmol·kg−1 BQ-610, with 20 nmol·kg−1 ETR-P1/fl (a novel ETA receptor antagonist) or with 20 nmol·kg−1 IRL 1038 (an ETB receptor antagonist).
The results of the separate assessments of the airway and tissue mechanics demonstrate that endothelin-1 induces airway and parenchymal constriction via stimulation of both receptor types in both compartments.
- airway resistance
- endothelin-1
- endothelin receptor antagonists
- lung elastance
- lung impedance
- parenchymal resistance
This study was supported by Hungarian Scientific Research Fund grants T30670 and T023089.
Endothelin-1 (ET-1) is a powerful regulator of smooth muscle tone in many systems in both physiological and pathophysiological conditions. ET-1 induces vasoconstrictor and bronchoconstrictor responses, which suggests that an upregulated ET-1 release may play an important role in a variety of pulmonary disorders, such as pulmonary hypertension or bronchial asthma. Vasoconstriction is mediated by the ETA receptors, whereas the ETB receptors mediate both vasoconstriction and vasodilation, depending on the species and the particular circulatory region 1. Inthe airways, the ETA and ETB receptor types coexist 2, 3 and both are involved in ET-1-induced bronchoconstriction 4, 5. In the guinea-pig, the ETB receptors predominate in the lung 3, and the proportion of ETB is greater in the bronchi than in the trachea 2.
The bronchoconstrictor nature of ET-1 has been concluded from the global changes in the mechanical parameters of the lung 6–11. However, the situation is confused by observations that pulmonary parenchymal tissues may also contribute to the mechanical alterations in the lung during bronchoconstriction 12–14, as demonstrated by recent studies involving a model-based evaluation of pulmonary impedance (ZL) 15–17.
In vivo evidence for ET-1-induced parenchymal changes is difficult to establish. Although Nagase etal. 5 observed that ET-1 may cause increases in airway and tissue resistance and tissue elastance in the guinea-pig lung, the contribution of the parenchyma to the net effects has not been fully elucidated. These authors measured inhomogeneous local alveolar pressures, and it has been demonstrated that such sampling may not be representative of the global state, since the regional variability of isolated mechanical alterations cannot be excluded 15. Separate characterization of the airway and parenchymal components has recently been achieved with the ZL-assessment technique, and it has been demonstrated that increasing doses of ET-1 elicit dose-dependent increases in the airway and parenchymal parameters in guinea-pigs 17.
The aim of the present study was to assess whether pretreatment with ET receptor antagonists would influence the pulmonary responses to two doses of exogenous ET-1. In a previous paper 17 it was demonstrated that increasing doses of ET-1 in the range 0.05–0.8 nmol·kg−1 induced dose-dependent elevations in the airway and parenchymal parameters in guinea-pigs. In the present study, two of these doses were chosen: the lowest dose (0.05 nmol·kg−1), in the previous study, caused slight but significant changes in the airway resistance, tissue damping and elastance, though still without signs of inhomogeneous constriction, whereas the higher dose (0.2 nmol·kg−1) was selected because this was previously observed to elicit significant paradox decreases in airway inertance, indicating an inhomogeneous peripheral airway constriction. As the involvement of the ETA and ETB receptors in the pulmonary mechanical responses to ET-1 is not clearly known, the aim was to study the roles of the two types of receptors in the changes elicited by ET-1 in the airways and lung parenchyma. Pretreatment with two types of ETA receptor antagonists was applied: BQ-610, a compound with specific ETA receptor antagonist properties 18, and ETR-P1/fl,a peptide with potent anti-ETA receptor activity in the cardiovascular system 19. IRL 1038, a selective ETB receptor antagonist 20 was also administered.
Material and methods
Animal preparation
The study was approved by the University Ethical Committee for the Protection of Animals in Scientific Research and performed in adherence to the National Institute of Health guidelines on the use of experimental animals.
The experiments were carried out in guinea-pigs (weighing 380–600 g) anaesthetized with pentobarbital sodium (30 mg·kg−1 i.p.). The animals were placed on a heating pad to maintain body temperature at 37°C. The right carotid artery and left jugular vein were cannulated for the monitoring of mean arterial blood pressure and drug administration, respectively. A plastic cannula (length=30 mm, internal diameter=2 mm) was inserted into the trachea and connected to a small animal respirator (Harvard App. Inc., South Natick, MA, USA) delivering a tidal volume of 6 mL·kg−1 at a frequency of 70·min−1. Following midline sternotomy, the chest was widely opened and a positive end-expiratory pressure of 2.5 cmH2O was applied. The guinea-pigs were paralysed with pipecuronium bromide (0.2 mg·kg−1). Additional doses of pentobarbital sodium (10 mg·kg−1) and pipecuronium bromide (0.05 mg·kg−1) were given as needed to maintain anaesthesia and paralysis, respectively.
Experimental protocol
After the surgical procedure, the animals were allowed to stabilize for 20 min. They were randomized to one or other of the five groups and each was given ET-1 (Alexis Corp., Läufelfingen, Switzerland) i.v. in two successive doses, the first dose at zero time (ET-11, 0.05 nmol·kg−1) and the second dose 10 min thereafter (ET-12, 0.2 nmol·kg−1). The following groups were studied: group ET-1 (n=9): only ET-1 doses; group BQ-61020 (n=8): pretreatment with 20 nmol·kg−1 BQ-610 (Alexis Corp., Läufelfingen, Switzerland); group BQ-61080 (n=6): pretreatment with 80 nmol·kg−1 BQ-610; group ETR-P1/fl (n=5), pretreatment with 20 nmol·kg−1 ETR-P1/fl (Kurabo Ltd, Osaka, Japan); and group IRL 1038 (n=9): pretreatment with 20 nmol·kg−1 IRL 1038 (Alexis Corp., Läufelfingen, Switzerland). The antagonists were administered 3 min prior to the first ET-1 dose (ET-11).
Pulmonary mechanics
Small-amplitude pseudorandom oscillations 0.5–21 Hz were introduced into the trachea for 6-s intervals at end-expiration to determine the ZL spectra as previously described 16, 17. Four measurements of ZL were made under the control conditions (before antagonist pretreatment and/or the administration of ET-11 and ET-12). Further ZL measurements were made at 0.5, 1, 2, 4 and 6 min after administration of each of the successive ET-1 boluses. Prior to the control ZL measurements, the lungs were hyperinflated by occluding the expiratory line of the respirator for two consecutive cycles to prevent atelectasis. This manoeuvre was repeated at 7 min after each ET-1 dose. A model containing an airway and a constant-phase tissue compartment was fitted to the ZL data 15; the airways were characterized by frequency-independent airway resistance (Raw) and inertance (Iaw), and the tissue properties were represented by coefficients of parenchymal damping (G) and elastance (H). Parenchymal hysteresivity (η) was calculated as G/H 21. The control ZL spectra and those recorded after ET-1 antagonist administration were averaged separately and then fitted, whereas the ZL data obtained following the administration of the two successive doses of ET-1 were fitted individually. The constrictor responses to each of the ET-1 doses were characterized by the parameter values obtained from ZL recorded at 0.5 min after administration. Before the experiments, the resistance and inertance of the tracheal cannula were also measured, and were subtracted from the corresponding airway resistance (Raw) and inertance (Iaw).
Gravimetric and morphometric analysis
At the end of each experiment, the animals were killed with an overdose of pentobarbital sodium, and lung tissue samples were taken for gravimetric and histological analysis. The wet weight (WW) of each sample was measured and the samples were then dried until a constant dry weight (DW) was reached. The wet-to-dry ratio (WW/DW) for each sample was calculated. Histological analysis was also performed. Tissue samples of removed lungs for routine histology were fixed in 10% neutral-buffered formalin and paraffin embedded. Sections of each sampled tissue were stained with haematoxylin and eosin and periodic acid-Schiff. The alveoli (oedema fluid, polymorphonuclear cells, macrophages) and the alveolar capillaries (congestion) were observed.
Statistical analysis
The results are presented as mean±sem both for the absolute values and for the percentage changes from the control data. Nonparametric tests were used forstatistical analysis. Friedman repeated measures analysis of variance on ranks with the Student-Newman-Keuls multiple comparison procedure, and Kruskal-Wallis one-way analysis of variance on ranks with the Dunn multiple comparison procedure, were used for within-group and between-group comparisons, respectively. A p-value of <0.05 was considered statistically significant.
Results
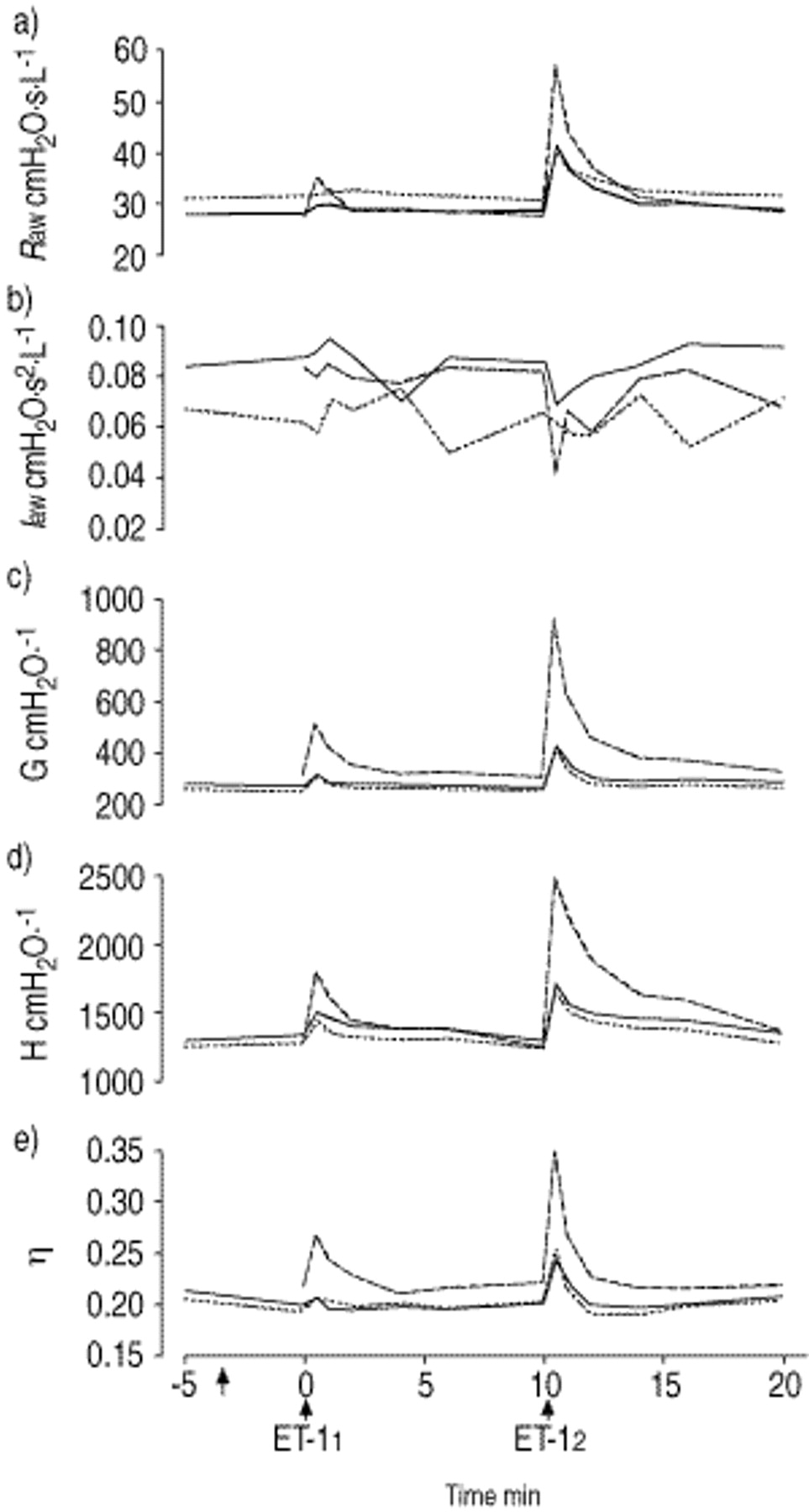
Figure 1⇓ presents the characteristic profiles of the changes in airway and parenchymal parameters after each of the two successive doses of ET-1, with and without pretreatment with the higher dose of BQ-610 or IRL 1038. There were only slight differences in baseline values between the individual groups; however, the control G value for group BQ-61020 was significantly different from that for group ETR-P1/fl. Both the first and the second doses of ET-1 induced significant increases in Raw, G, H and η in group ET-1, and this dose-dependent pattern remained in all groups which received antagonists. The parameters reached their maxima within 30 s after administration of either dose of ET-1, and the peak values then gradually declined toward the baseline. In group BQ-61020, the elevations in respiratory parameters were close to those observed in group ET-1. However, in groups BQ-61080, IRL 1038 and ETR-P1/fl, each of the doses of ET-1 evoked significantly smaller increases in Raw, G, H and η. The patterns of change in Iaw were inconsequential: in all groups but group BQ-61020, no statistically significant changes in Iaw were observed after either ET-1 dose.

Mean a) airway resistance (Raw); b) airway inertance (Iaw), c) tissue damping (G), d) elastance (H) and e) G/H ratio (η) parameter profiles induced by two successive doses of (0 and 10 min; ↑) of endothelin-1 (ET-11 and ET-12), with and without pretreatment (↑) with one or other of two endothelin receptor antagonists. – – –: ET-1 treatment only; - - -: ET-1 treatment after pretreatment with BQ-61080; —: ET-1 treatment after pretreatment with IRL 1038.
In group ET-1, the durations of significant changes in Raw and H were dose-dependent. Significant changes in η persisted for 1 min, and those in G for 10 min after each of the ET-1 doses (table 1⇓). The lower dose of BQ-610 significantly shortened the duration of changes in G, and increased those in η after ET-11. The responses in Raw, G and η were statistically significant only for short intervals in the cases of the higher dose of BQ-610, IRL 1038 and ETR-P1/fl.
Mean durations (min) of significant (P<0.05) increases in airway and parenchymal parameters induced by two successive endotheliun-1 (ET-1) doses (ET-11 and ET-12), without (group ET-1) and with pretreatment with an ET-1 receptor antagonist
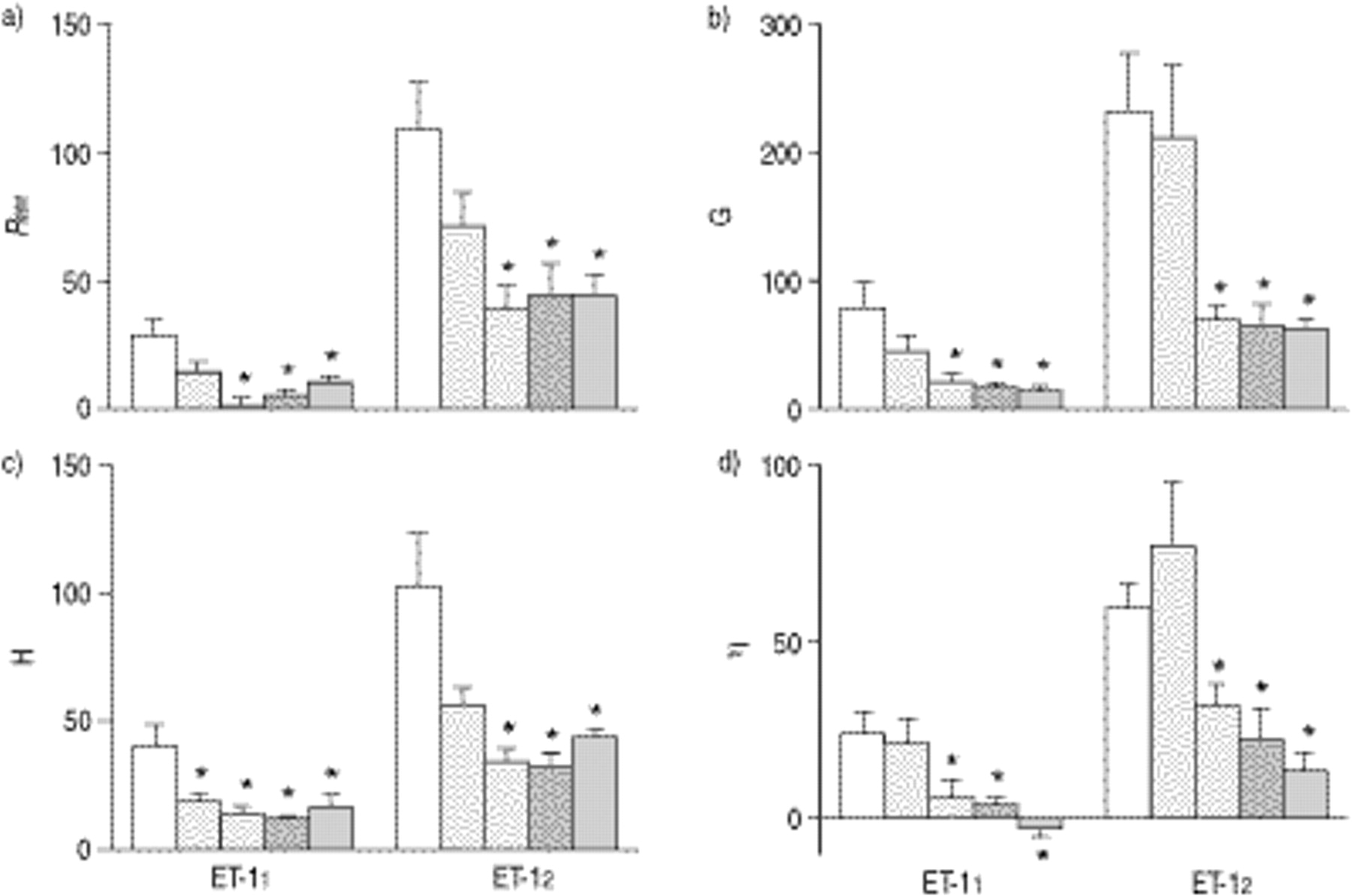
Because of the high variations in the baseline data, the percentage changes in the airway (Raw) and parenchymal parameters (G, H and η) were calculated by normalizing the values measured at 30 s after each of the ET-1 doses to those obtained before ET-1 administration (fig. 2⇓). ET-1 induced dose-dependent increases in each of these parameters in group ET-1. In group BQ-61020 after ET-11, the differences from the corresponding values in group ET-1 reached the level of statistical significance only for H. The responses in Raw, G, H and η after each ET-1 dose were significantly smaller in groups BQ-61080, IRL 1038 and ETR-P1/fl as compared with those in group ET-1.

{kind=link}
{kind=link}
Changes induced in the parameters a) airway resistance (Raw), b) tissue damping (G), elastance (H) and G/H ratio (η) by two successive endothelin-1 (ET-1) doses (ET-11 and ET-12) with and without pretreatment with an endothelin receptor antagonist. The parameters were measured at 30 s after each ET-1 dose and are normalized to the control values measured before ET-1 administration. *: p<0.05 versus the value for ET-1. □: ET-1 treatments only; ┘: ET-1 and BQ-61020 treatment; └: ET-1 and BQ61080 treatment;  : ET-1 and IRL 1038 treatment;
: ET-1 and IRL 1038 treatment;  : ET-1 and ETR-P1/fl treatment.
: ET-1 and ETR-P1/fl treatment.
No significant differences in the counts of polymorphonuclear cells and macrophages between the groups were obtained; in addition, no signs of alveolar oedema were observed (data not shown). The absence of oedema was supported by the results of gravimetric analysis of the tissue samples. The ratios WW/DW in the antagonist-treated groups did not differ significantly from that for group ET-1 (ET-1: 5.11±0.1; BQ-61020: 4.88±0.07; BQ-61080: 5.63±0.2; IRL 1038: 5.06±0.1; and ETR-P1/fl: 5.19±0.1).
Discussion
This study confirms that exogenous ET-1 induces significant changes in the mechanical properties of the airways and parenchyma in the guinea-pig lung. Administration of ETA and ETB receptor antagonists alone did not change the basal values of the mechanical parameters, which suggests that endogenous ET-1 does not play a significant role in the regulation of the resting bronchomotor tone in the guinea-pig. However, each of the doses of ET-1 significantly increased Raw, G, H and η. The lower dose of BQ-610 significantly decreased only the changes in H after ET-11, whereas the higher dose of the antagonist also reduced those in Raw, G, and η. Pretreatment with IRL 1038 and ETR-P1/fl reduced the responses to ET-1 in both the airways and the parenchyma. Thus, it is concluded that ET-1 induces receptor-sensitive parenchymal reactions that are distinct from the airway responses.
Endothelin-1 receptor antagonists
The selective ETA receptor antagonist BQ-610, the ETA receptor antagonist ETR-P1/fl and the selective ETB receptor antagonist IRL 1038 were used to investigate the roles of the ET receptors in the mechanical properties of the airways and the parenchyma. BQ-610 is a highly selective ETA receptor antagonist, being an ∼30,000 times more effective inhibitor of ETA than of ETB receptors in vitro 18, and it effectively antagonizes the in vivo effects of ET-1 on the ETA receptors 22, 23. ETR-P1/fl is an “antisense-homology box”-derived peptide, which exhibits anti-ETA receptor activity both in vitro 19 and in vivo 22, 23. Additionally, it improves the symptoms and haemodynamic parameters in endotoxin shock 24 and in hypodynamic septic rats 23, and inhibits the endothelial cell-leukocyte interactions in response to ET-1 in the intestinal microcirculation in rats 22. IRL 1038 binds specifically to the ETB receptors in various mammalian tissues, including the guinea-pig lung 20.
Methodological view
Before comparing the results with other data in the literature, it is relevant to discuss certain aspects of the methodology. It has previously been demonstrated that i.v. ET-1 induces dose-dependent increases in the peak pulmonary inflation pressure 8–10 or elevations in total lung resistance and decreases in dynamic lung compliance 6, 7, 11. However, it should be stressed that these parameters are indicators only of the changes in the global lung mechanics: the peak inflation pressure, combining the resistive and elastic responses, and the total respiratory or pulmonary resistance, combining the airway and tissue properties in a frequency-dependent manner. The present technique, involving a model-based evaluation of multiple-frequency ZL data, allows estimation of the parameters that characterize the airway and parenchymal mechanics separately, both under control conditions and during constrictions induced by drugs 15, 16, 25, including ET-1 17. In the latter study, the mechanical responses were similar at corresponding doses of ET-1. There are no data that are directly comparable with the present findings, and only one paper reports parenchymal changes induced by ET-1 in the guinea-pig invivo 5.
Airway and parenchymal responses
In the present study, the two successive doses of ET-1 each caused significant dose-related changes in the mechanical properties of the airways and the parenchyma. Of the airway parameters, Raw was increased significantly, without any significant change in Iaw. Since Iaw is dominated by the inertance in the central airways, it is concluded that ET-1 induces airway constriction in the periphery of the lung. IRL 1038, ETR-P1/fl and the higher dose of BQ-610 significantly decreased the elevation induced in Raw by each of the ET-1 doses.
Autoradiographic studies have demonstrated that ETA and ETB receptors coexist in the guinea-pig lung, with an ETA:ETB receptor ratio of 1:4 3. Furthermore, Hay et al. 2 described regional differences in the distributions of the two ET receptor types in the airways. The primary bronchi contain more ETB receptors, whereas the ETA receptors predominate in the trachea. The relative contribution of the ETB receptors to bronchoconstriction, therefore, increases in the distal direction along the respiratory tract 2. Pharmacological investigations in isolated guinea-pig bronchi provided evidence that ET-1 causes bronchoconstriction largely via ETB receptor stimulation 4, 26. Nagase et al. 5 found that both ETA andETB receptor antagonists effectively reduce the ET-1-induced increases in airway resistance in vivo. In the present experiments, IRL 1038, ETR-p1/fl and the higher dose of BQ-610 significantly decreased the elevation in Raw after ET-1 administration. These data, therefore, indicate that both types of receptors are involved in the ET-1-induced increase in Raw.
The present study found that ET-1 produced significant elevations in the parenchymal parameters G, H and η, indicating substantial changes in the mechanical properties of the parenchyma. Pretreatment with IRL 1038, ETR-P1/fl and the higher dose of BQ-610 significantly diminished the responses in H after each of the ET-1 doses. Nagase et al. 5 reported that the responses in tissue resistance and lung elastance to ET-1 did not differ significantly in the presence of BQ-123, whereas BQ-788 significantly decreased the responses in both tissue parameters. Battistini et al. 4 demonstrated that BQ-123 inhibited the ET-1-induced contraction in parenchymal strips in vitro; however, this treatment was ineffective in the bronchi. In the present experiments, pretreatment with either dose of BQ-610, ETR-P1/fl or IRL 1038 significantly reduced the changes in H. These findings suggest that both types of ET receptors are involved in themediation of ET-1-induced constriction in the parenchyma.
It has been proposed that a heterogeneous population of ETB receptors exists in the guinea-pig respiratory system 4, 9, 27. In the present experiments, pretreatment with BQ-610 in a dose of 20 nmol·kg−1 had practically no effect on the ET-1-induced airway and parenchymal responses. However, pretreatment with the higher dose significantly decreased the changes in the airway and parenchymal parameters, suggesting that the higher concentration of this ETA antagonist resulted in the inhibition of both ETA and ETB receptors. The dose-dependent effect of the ETA receptor antagonist BQ-610 on the ET-1-induced pulmonary constriction provides additional evidence for the hypothesis that there may be an ETA receptor antagonist-binding ETB receptor subtype in the guinea-pig lung 4, 9, 27.
ETR-P1/fl peptide
The present study provides the first demonstration of the effects of the ETA receptor antagonist ETR-P1/fl on the ET-1-induced mechanical changes in the lung. In the presence of ETR-P1/fl, significantly smaller elevations in Raw, G, H and η occurred after each dose of ET-1 than those observed in untreated animals. Moreover, the peptide decreased the durations of the significant changes in Raw, G and η after each ET-1 dose. Thus, ETR-P1/fl affected the pulmonary mechanics in the same dose as did IRL 1038, but not BQ-610. Therefore, it is reasonable to suggest that ETR-P1/fl is not selective for ETA receptors in the guinea-pig lung. These findings, in a similar manner to those of Szalay et al. 23, indicate that ETR-P1/fl may antagonize not only ETA, but also ETB receptors. Another explanation may be that ETR-P1/fl removes circulating ET-1, as was observed by Baranyi et al. 24 in dogs.
Gravimetric and morphometric results
ET-1 has been shown to be a pro-inflammatory mediator. It induces intravascular leukocyte sequestration, and enhances albumin extravasation and oedema formation in the guinea-pig lung 28. Filep et al. 28 found that ET-1 in doses of 0.3 and 1 nmol·kg−1 gives rise to albumin extravasation via activation of the ETA receptors in the trachea and the bronchi, but not in the parenchyma. However, the present study shows that ET-1 doses of 0.05 and 0.2 nmol·kg−1 do not elicit lung oedema, as evidenced by gravimetric and histological examinations. In agreement with these data, Macquin-Mavier et al. 6 did not observe significant lung oedema in ET-1-treated animals. At present, an explanation cannot be given for these discordant results, and the dynamics of ET-1-induced oedema formation should be further investigated.
Secondary mediators
Several studies have established that various mediators can contribute to the mechanical changes induced by ET-1 in the lung. It has been demonstrated that the bronchoconstriction evoked by ET-1 could be mediated by thromboxane A2 released via ET receptor activation 8, 29, 30. Further, in the guinea-pig lung ET-1 can release the bronchodilator prostacyclin 30 and nitric oxide 29. The direct bronchoconstrictor effect of ET-1 can, therefore, be either potentiated or counteracted by secondary mediators.
In summary, this study has characterized the pulmonary mechanical responses to the administration of endothelin-1, both with and without pretreatment with ETA and ETB receptor antagonists, with a method appropriate for the separate quantification of the airway and parenchymal components. The results reveal that the activation of both ETA and ETB receptor subtypes is involved in the mediation of the mechanical changes in response to endothelin-1 in the airways and the parenchyma.
Acknowledgments
The authors are grateful to H. Okada and L. Baranyi for their generous supply of the ETR-P1/fl peptide, and to Á. Fekete, A. Nagyiván, I. Kopasz and L. Vígh for their skilful technical assistance.
- Received March 16, 2000.
- Accepted January 19, 2001.
- © ERS Journals Ltd
References









